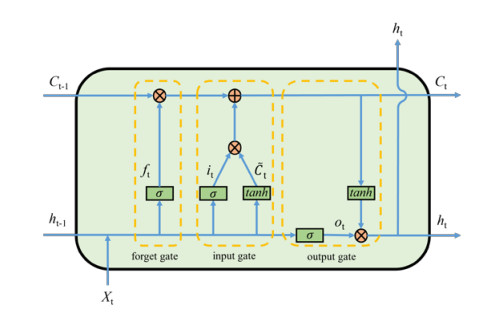
There is limited research on the loss and reconstruction of car-following features. To delve into car-following's characteristics, we propose a car-following model based on LSTM-Transformer. By fully leveraging the advantages of long short-term memory (LSTM) and transformer models, this study focuses on reconstructing the input car-following features. Training and testing were conducted using 700 car-following segments extracted from a natural driving dataset and the Next Generation Simulation (NGSIM) dataset, and the proposed model was compared with an LSTM model and an intelligent driver model. The results demonstrate that the model performs exceptionally well in feature reconstruction. Moreover, compared to the other two models, it effectively captures the car-following features and accurately predicts the position and speed of the following car when features are lost. Additionally, the LSTM-Transformer model accurately reproduces traffic phenomena, such as asymmetric driving behavior, traffic oscillations and lag, by reconstructing the lost features. Therefore, the LSTM-Transformer car-following model proposed in this study exhibits advantages in feature reconstruction and reproducing traffic phenomena compared to other models.
Citation: Pinpin Qin, Xing Li, Shenglin Bin, Fumao Wu, Yanzhi Pang. Research on transformer and long short-term memory neural network car-following model considering data loss[J]. Mathematical Biosciences and Engineering, 2023, 20(11): 19617-19635. doi: 10.3934/mbe.2023869
There is limited research on the loss and reconstruction of car-following features. To delve into car-following's characteristics, we propose a car-following model based on LSTM-Transformer. By fully leveraging the advantages of long short-term memory (LSTM) and transformer models, this study focuses on reconstructing the input car-following features. Training and testing were conducted using 700 car-following segments extracted from a natural driving dataset and the Next Generation Simulation (NGSIM) dataset, and the proposed model was compared with an LSTM model and an intelligent driver model. The results demonstrate that the model performs exceptionally well in feature reconstruction. Moreover, compared to the other two models, it effectively captures the car-following features and accurately predicts the position and speed of the following car when features are lost. Additionally, the LSTM-Transformer model accurately reproduces traffic phenomena, such as asymmetric driving behavior, traffic oscillations and lag, by reconstructing the lost features. Therefore, the LSTM-Transformer car-following model proposed in this study exhibits advantages in feature reconstruction and reproducing traffic phenomena compared to other models.

| [1] |
V. Papathanasopoulou, C. Antoniou, Towards data-driven car-following models, Transp. Res. Part C Emerging Technol., 55 (2015), 496–509. https://doi.org/10.1016/j.trc.2015.02.016 doi: 10.1016/j.trc.2015.02.016

|
| [2] |
M. Saifuzzaman, Z. Zheng, Incorporating human-factors in car-following models: A review of recent developments and research needs, Transp. Res. Part C Emerging Technol., 48 (2014), 379–403. https://doi.org/10.1016/j.trc.2014.09.008 doi: 10.1016/j.trc.2014.09.008
|
| [3] |
V. Punzo, Z. Zheng, M. Montanino, About calibration of car-following dynamics of automated and human-driven vehicles: Methodology, guidelines and codes, Transp. Res. Part C Emerging Technol., 128 (2021), 103165. https://doi.org/10.1016/j.trc.2021.103165 doi: 10.1016/j.trc.2021.103165
|
| [4] |
Z. Mo, R. Shi, X. Di, A physics-informed deep learning paradigm for car-following models, Transp. Res. Part C Emerging Technol., 130 (2021), 103240. https://doi.org/10.1016/j.trc.2021.103240 doi: 10.1016/j.trc.2021.103240
|
| [5] |
J. Liu, R. Jiang, J. Zhao, W. Shen, A quantile-regression physics-informed deep learning for car-following model, Transp. Res. Part C Emerging Technol., 154 (2023), 104275. https://doi.org/10.1016/j.trc.2023.104275 doi: 10.1016/j.trc.2023.104275
|
| [6] | Z. Mo, X. Di, Uncertainty quantification of car-following behaviors: physics-informed generative adversarial networks, the 28th ACM SIGKDD in conjunction with the 11th International Workshop on Urban Computing (UrbComp2022), 2022. Available from: http://urban-computing.com/urbcomp2022/file/UrbComp2022_paper_3574. |
| [7] |
M. Montanino, V. Punzo, Trajectory data reconstruction and simulation-based validation against macroscopic traffic patterns, Transp. Res. Part B Methodol., 80 (2015), 82–106. https://doi.org/10.1016/j.trb.2015.06.010 doi: 10.1016/j.trb.2015.06.010
|
| [8] |
X. Wang, Y. Ge, L. Niu, Y. He, T. Z. Qiu, Method for imputing missing data using online calibration for urban freeway control, Transp. Res. Rec., 2672 (2018), 44–54. https://doi.org/10.1177/0361198118774735 doi: 10.1177/0361198118774735
|
| [9] |
S. Tak, S. Woo, H. Yeo, Data-driven imputation method for traffic data in sectional units of road links, IEEE Trans. Intell. Transp. Syst., 17 (2016), 1762–1771. https://doi.org/10.1109/TITS.2016.2530312 doi: 10.1109/TITS.2016.2530312
|
| [10] |
J. M. Chiou, Y. C. Zhang, W. H. Chen, C. W. Chang, A functional data approach to missing value imputation and outlier detection for traffic flow data, Transportmetrica B: Transport Dyn., 2 (2014), 106–129. https://doi.org/10.1080/21680566.2014.892847 doi: 10.1080/21680566.2014.892847
|
| [11] |
J. Tang, Y. Wang, S. Zhang, H. Wang, F. Liu, S. Yu, On missing traffic data imputation based on fuzzy C-means method by considering spatial–temporal correlation, Transp. Res. Rec., 2528 (2019), 86–95. https://doi.org/10.3141/2528-10 doi: 10.3141/2528-10
|
| [12] |
J. Zhao, Y. Gao, J. Tang, L. Zhu, J. Ma, Highway travel time prediction using sparse tensor completion tactics and K-Nearest neighbor pattern matching method, J. Adv. Transp., 2018 (2018), 1–16. https://doi.org/10.1155/2018/5721058 doi: 10.1155/2018/5721058
|
| [13] |
Y. Duan, Y. Lv, Y. L. Liu, F. Y. Wang, An efficient realization of deep learning for traffic data imputation, Transp. Res. Part C Emerging Technol., 72 (2016), 168–181. https://doi.org/10.1016/j.trc.2016.09.015 doi: 10.1016/j.trc.2016.09.015
|
| [14] |
Y. Zhuang, R. Ke, Y. Wang, Innovative method for traffic data imputation based on convolutional neural network, IET Intell. Transp. Syst., 13 (2018), 605–613. https://doi.org/10.1049/iet-its.2018.5114 doi: 10.1049/iet-its.2018.5114
|
| [15] |
D. Zhao, Y. Zhang, W. Wang, X. Hua, M. Yang, Car‐following trajectory data imputation with adversarial convolutional neural network, IET Intell. Transp. Syst., 17 (2022), 960–972. https://doi.org/10.1049/itr2.12319 doi: 10.1049/itr2.12319
|
| [16] |
Y. Liang, Z. Zhao, L. Sun, Memory-augmented dynamic graph convolution networks for traffic data imputation with diverse missing patterns, Transp. Res. Part C Emerging Technol., 143 (2022), 103826. https://doi.org/10.1016/j.trc.2022.103826 doi: 10.1016/j.trc.2022.103826
|
| [17] |
C. Zhao, A. Song, Y. Du, B. Yang, TrajGAT: A map-embedded graph attention network for real-time vehicle trajectory imputation of roadside perception, Transp. Res. Part C Emerging Technol., 142 (2022), 103787. https://doi.org/10.1016/j.trc.2022.103787 doi: 10.1016/j.trc.2022.103787
|
| [18] |
Q. Wan, G. Peng, Z. Li, F. H. T. Inomata, Spatiotemporal trajectory characteristic analysis for traffic state transition prediction near expressway merge bottleneck, Transp. Res. Part C Emerging Technol., 117 (2020), 102682. https://doi.org/10.1016/j.trc.2020.102682 doi: 10.1016/j.trc.2020.102682
|
| [19] |
A. Sherstinsky, Fundamentals of Recurrent Neural Network (RNN) and Long Short-TermMemory (LSTM) network, Physica D, 404 (2020), 132306. https://doi.org/10.1016/j.physd.2019.132306 doi: 10.1016/j.physd.2019.132306
|
| [20] |
S. Dong, P. Wang, K. Abbas, A survey on deep learning and its applications, Comput. Sci. Rev., 40 (2021), 100379. https://doi.org/10.1016/j.cosrev.2021.100379 doi: 10.1016/j.cosrev.2021.100379
|
| [21] |
C. Sun, J. Leng, F. Sun, A fast optimal speed planning system in arterial roads for intelligent and connected vehicles, IEEE Internet Things J., 9 (2022), 20295–20307. https://doi.org/10.1109/JIOT.2022.3172009 doi: 10.1109/JIOT.2022.3172009
|
| [22] |
P. Qin, H. Li, Z. Li, W. Guan, Y. He, A CNN-LSTM car-following model considering generalization ability, Sensors, 23 (2023), 660. https://doi.org/10.3390/s23020660 doi: 10.3390/s23020660
|
| [23] |
L. Ma, S. Qu, J. Ren, X. Zhang, Mixed traffic flow of human-driven vehicles and connected autonomous vehicles: String stability and fundamental diagram, Math. Biosci. Eng., 20 (2022), 2280–2295. https://doi.org/10.3934/mbe.2023107 doi: 10.3934/mbe.2023107
|
| [24] |
L. Qu, L. Li, Y. Zhang, J. Hu, PPCA-based missing data imputation for traffic flow volume: A systematical approach, IEEE Trans. Intell. Transp. Syst., 10 (2009), 512–522. https://doi.org/10.1109/TITS.2009.2026312 doi: 10.1109/TITS.2009.2026312
|
| [25] | T. W. Forbes, Human factor considerations in traffic flow theory, Highway Res. Rec., (1963), 60–66. |
| [26] |
H. Gong, H. Liu, B. H. Wang, An asymmetric full velocity difference car-following model, Physica A, 387 (2008), 2595–2602. https://doi.org/10.1016/j.physa.2008.01.038 doi: 10.1016/j.physa.2008.01.038
|
| [27] |
D. Wei, H. Liu, Analysis of asymmetric driving behavior using a self-learning approach, Transp. Res. Part B Methodol., 47 (2013), 1–14. https://doi.org/10.1016/j.trb.2012.09.003 doi: 10.1016/j.trb.2012.09.003
|
| [28] | G. F. Newell, Instability in dense highway traffic: A review, Highway Res. Rec., (1965), 73–83. |
| [29] | L. C. Edie, Discussion of Traffic Stream Measurements and Definitions, Port of New York Authority, (1965), 139–154. |
| [30] | J. Treiterer, J. Myers, The hysteresis phenomenon in traffic flow, Transp. Traffic Theory, 6 (1974), 13–38. |
| [31] |
J. A. Laval, Hysteresis in traffic flow revisited: An improved measurement method, Transp. Res. Part B Methodol., 45 (2011), 385–391. https://doi.org/10.1016/j.trb.2010.07.006 doi: 10.1016/j.trb.2010.07.006
|








Figures(9) / Tables(5)

Pinpin Qin, Xing Li, Shenglin Bin, Fumao Wu, Yanzhi Pang. Research on transformer and long short-term memory neural network car-following model considering data loss[J]. Mathematical Biosciences and Engineering, 2023, 20(11): 19617-19635. doi: 10.3934/mbe.2023869







 DownLoad:
DownLoad: