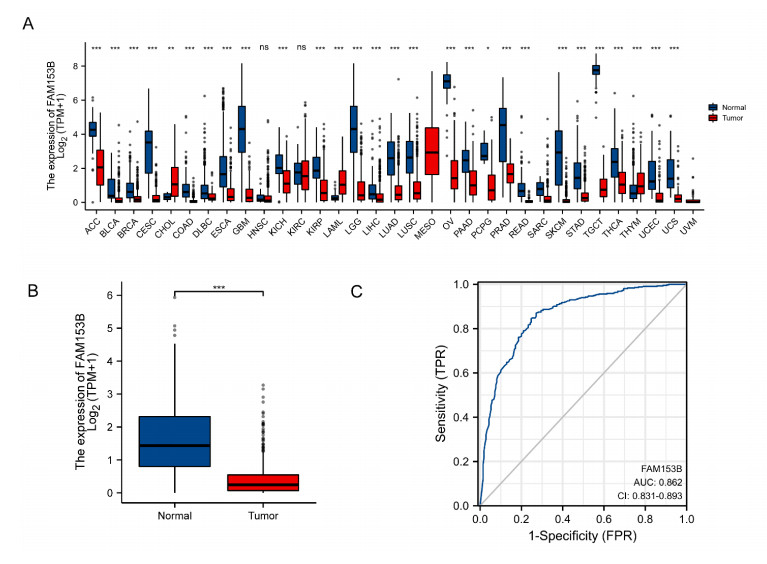
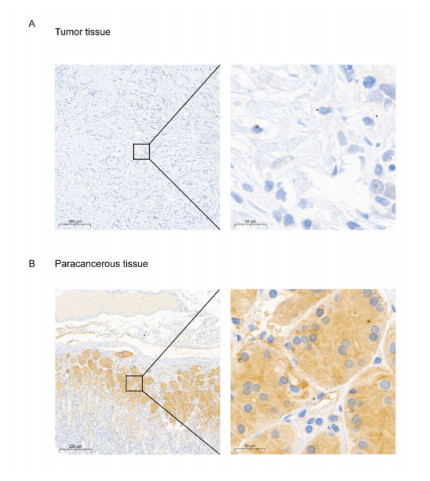
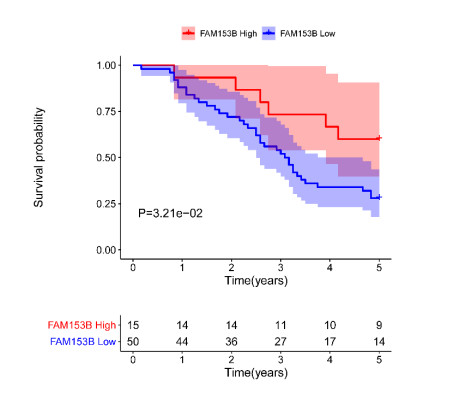
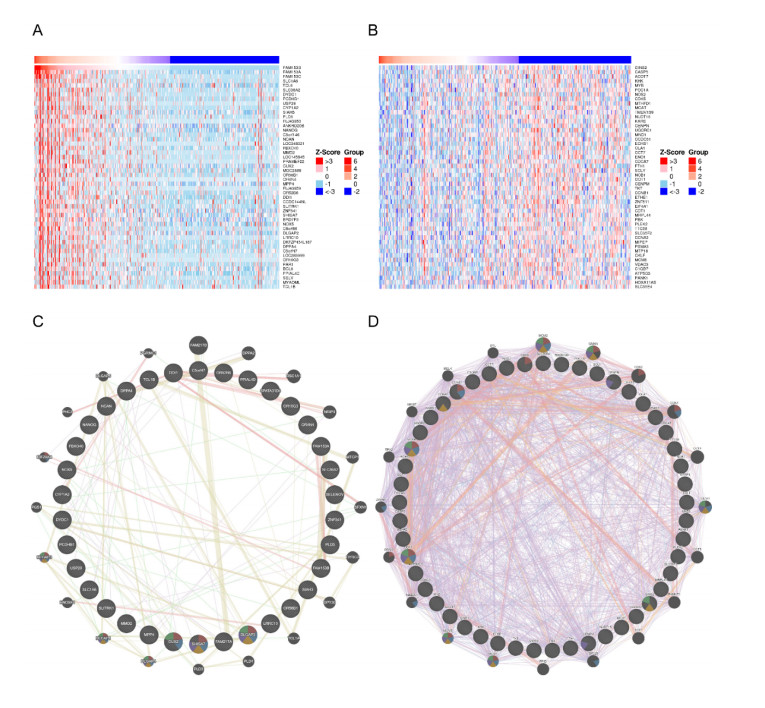
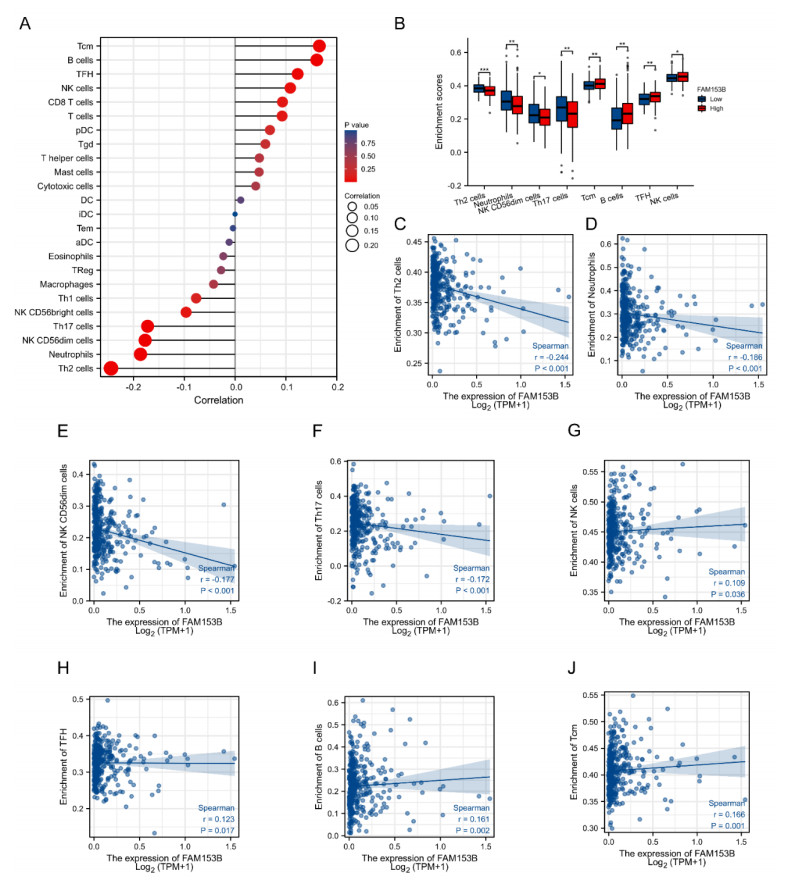
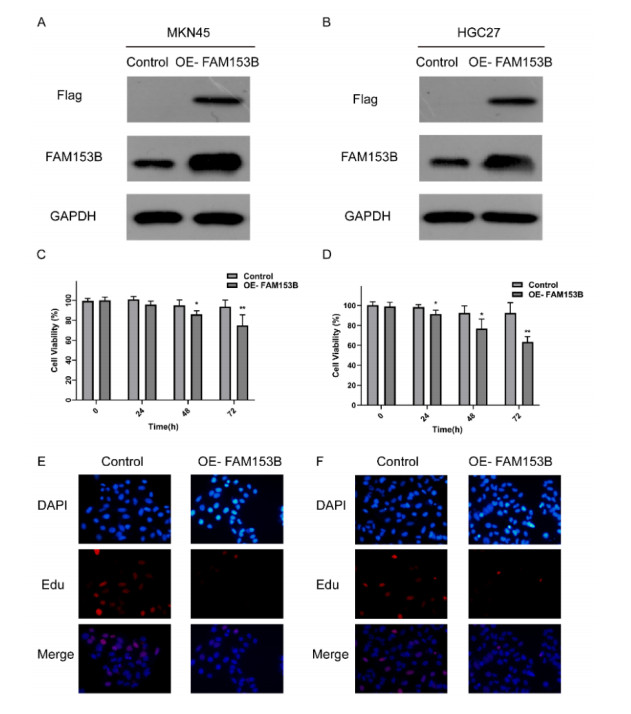
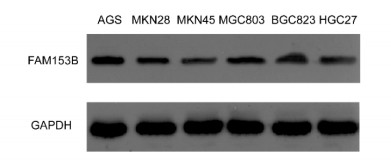
Gastric cancer (GC) is one of the most common digestive tumors in Northwest China. Previous sequencing analysis revealed that family with sequence similarity 153 member B (FAM153B) might be the primary driver gene of GC. In this study, we aim to explore the potential roles of FAM153B in GC. Microarray data were firstly obtained from public databases with the aim to evaluate the genetic expression of FAM153B between GC and normal tissues. The results were verified in immunohistochemistry (IHC). We also performed the co-expression network analysis and enrichment analysis to identify underlying mechanisms. A correlation analysis of FAM153B expression and immune infiltration was performed then. Furthermore, two GC cell lines were used to evaluate the effect of FAM153B on gastric cell proliferation by employing MTT and Edu assays. Our findings suggest that FAM153B is downregulated in tumoral tissue, and positively associated with unfavorable survival. The enrichment pathways of FAM153B were regulation of signaling receptor activity, DNA replication, cell cycle transition, chromosomal regulation, and so on. Besides, from the perspective of bioinformatics, the protein expression level of FAM153B is related to the degree of immune cell infiltration. In vitro, overexpression of FAM153B inhibit the proliferation of two cell lines. In summary, this study demonstrates that FAM153B might serve as an effective prognostic and therapeutic biomarker in GC.
Citation: Yuan Yang, Yuwei Ye, Min Liu, Ya Zheng, Guozhi Wu, Zhaofeng Chen, Yuping Wang, Qinghong Guo, Rui Ji, Yongning Zhou. Family with sequence similarity 153 member B as a potential prognostic biomarker of gastric cancer[J]. Mathematical Biosciences and Engineering, 2022, 19(12): 12581-12600. doi: 10.3934/mbe.2022587
Gastric cancer (GC) is one of the most common digestive tumors in Northwest China. Previous sequencing analysis revealed that family with sequence similarity 153 member B (FAM153B) might be the primary driver gene of GC. In this study, we aim to explore the potential roles of FAM153B in GC. Microarray data were firstly obtained from public databases with the aim to evaluate the genetic expression of FAM153B between GC and normal tissues. The results were verified in immunohistochemistry (IHC). We also performed the co-expression network analysis and enrichment analysis to identify underlying mechanisms. A correlation analysis of FAM153B expression and immune infiltration was performed then. Furthermore, two GC cell lines were used to evaluate the effect of FAM153B on gastric cell proliferation by employing MTT and Edu assays. Our findings suggest that FAM153B is downregulated in tumoral tissue, and positively associated with unfavorable survival. The enrichment pathways of FAM153B were regulation of signaling receptor activity, DNA replication, cell cycle transition, chromosomal regulation, and so on. Besides, from the perspective of bioinformatics, the protein expression level of FAM153B is related to the degree of immune cell infiltration. In vitro, overexpression of FAM153B inhibit the proliferation of two cell lines. In summary, this study demonstrates that FAM153B might serve as an effective prognostic and therapeutic biomarker in GC.

| [1] |
H. Sung, J. Ferlay, R. L. Siegel, M. Laversanne, I. Soerjomataram, A. Jemal, et al., Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries, CA: A Cancer J. Clin., 71 (2021), 209–249. https://doi.org/10.3322/caac.21660 doi: 10.3322/caac.21660

|
| [2] |
R. E. Sexton, M. N. Al Hallak, M. Diab, A. S. Azmi, Gastric cancer: a comprehensive review of current and future treatment strategies, Cancer Metastasis Rev., 39 (2020), 1179–1203. https://doi.org/10.1007/s10555-020-09925-3 doi: 10.1007/s10555-020-09925-3
|
| [3] |
M. Banks, D. Graham, M. Jansen, T. Gotoda, S. Coda, M. di Pietro, et al., British Society of Gastroenterology guidelines on the diagnosis and management of patients at risk of gastric adenocarcinoma, BMJ, 68 (2019), 1545–1575. https://doi.org/10.1136/gutjnl-2018-318126 doi: 10.1136/gutjnl-2018-318126
|
| [4] |
A. Zayac, K. Almhanna, Esophageal, gastric cancer and immunotherapy: small steps in the right direction, Transl. Gastroenterol. Hepatol., 5 (2020), 9. https://doi.org/10.21037/tgh.2019.09.05 doi: 10.21037/tgh.2019.09.05
|
| [5] |
A. P. Thrift, H. B. El-Serag, Burden of gastric cancer, Clin. Gastroenterol. Hepatol., 18 (2020), 534–542. https://doi.org/10.1016/j.cgh.2019.07.045 doi: 10.1016/j.cgh.2019.07.045
|
| [6] |
M. Cao, H. Li, D. Sun, W. Chen, Cancer burden of major cancers in China: a need for sustainable actions, Cancer Commun., 40 (2020), 205–210. https://doi.org/10.1002/cac2.12025 doi: 10.1002/cac2.12025
|
| [7] |
T. Matsuoka, M. Yashiro, Biomarkers of gastric cancer: current topics and future perspective, World J. Gastroenterol., 24 (2018), 2818–2832. https://doi.org/10.3748/wjg.v24.i26.2818 doi: 10.3748/wjg.v24.i26.2818
|
| [8] |
B. Ferdouse, I. Ruczinski, S. Li, E. Silverman, M. Cho, D. Lynch, et al., Identifying a deletion affecting total lung capacity among subjects in the COPDGene study cohort, Genet Epidemiol, 1 (2016), 81–88. https://doi.org/10.1002/gepi.21943 doi: 10.1002/gepi.21943
|
| [9] |
M. Macheret, T. D. Halazonetis, DNA replication stress as a hallmark of cancer, Annu. Rev. Pathol.: Pathol. Mech. Dis., 10 (2015), 425–448. https://doi.org/10.1146/annurev-pathol-012414-040424 doi: 10.1146/annurev-pathol-012414-040424
|
| [10] |
H. Zhu, U. Swami, R. Preet, J. Zhang, Harnessing DNA replication stress for novel cancer therapy, Genes, 11 (2020), 990. https://doi.org/10.3390/genes11090990 doi: 10.3390/genes11090990
|
| [11] |
H. Kitao, M. Iimori, Y. Kataoka, T. Wakasa, E. Tokunaga, H. Saeki, et al., DNA replication stress and cancer chemotherapy, Cancer Sci., 109 (2018), 264–271. https://doi.org/10.1111/cas.13455 doi: 10.1111/cas.13455
|
| [12] |
J. Shi, X. Zhang, J. Li, W. Huang, Y. Wang, Y. Wang, et al., MTA2 sensitizes gastric cancer cells to PARP inhibition by induction of DNA replication stress, Transl. Oncol., 14 (2021), 101167. https://doi.org/10.1016/j.tranon.2021.101167 doi: 10.1016/j.tranon.2021.101167
|
| [13] |
M. Bauer, Z. Nascakova, A. I. Mihai, P. F. Cheng, M. P. Levesque, S. Lampart, et al., The ALPK1/TIFA/NF-κB axis links a bacterial carcinogen to R-loop-induced replication stress, Nat. Commun., 11 (2020), 5117. https://doi.org/10.1038/s41467-020-18857-z doi: 10.1038/s41467-020-18857-z
|
| [14] |
N. S. Sethi, O. Kikuchi, G. N. Duronio, M. D. Stachler, J. M. McFarland, R. Ferrer-Luna, et al., Early TP53 alterations engage environmental exposures to promote gastric premalignancy in an integrative mouse model, Nat. Genet., 52 (2020), 219–230. https://doi.org/10.1038/s41588-019-0574-9 doi: 10.1038/s41588-019-0574-9
|
| [15] |
J. Rozacky, A. A. Nemec, J. B. Sweasy, D. Kidane, Gastric cancer associated variant of DNA polymerase beta (Leu22Pro) promotes DNA replication associated double strand breaks, Oncotarget, 6 (2015), 24474–24487. https://doi.org/10.18632/oncotarget.4426 doi: 10.18632/oncotarget.4426
|
| [16] |
H. Hua, S. E. Kearsey, Monitoring DNA replication in fission yeast by incorporation of 5-ethynyl-2'-deoxyuridine, Nucleic Acids Res., 39 (2011), e60. https://doi.org/10.1093/nar/gkr063 doi: 10.1093/nar/gkr063
|
| [17] |
E. R. Mardis, Neoantigens and genome instability: impact on immunogenomic phenotypes and immunotherapy response, Genome Med., 11 (2019), 71. https://doi.org/10.1186/s13073-019-0684-0 doi: 10.1186/s13073-019-0684-0
|
| [18] |
D. Bruni, H. K. Angell, J. Galon, The immune contexture and immunoscore in cancer prognosis and therapeutic efficacy, Nat. Rev. Cancer, 20 (2020), 662–680. https://doi.org/10.1038/s41568-020-0285-7 doi: 10.1038/s41568-020-0285-7
|
| [19] |
W. H. Fridman, L. Zitvogel, C. Sautès-Fridman, G. Kroemer, The immune contexture in cancer prognosis and treatment, Nat. Rev. Clin. Oncol., 14 (2017), 717–734. https://doi.org/10.1038/nrclinonc.2017.101 doi: 10.1038/nrclinonc.2017.101
|
| [20] |
S. Y. Gun, S. W. L. Lee, J. L. Sieow, S. C. Wong, Targeting immune cells for cancer therapy, Redox Biol., 25 (2019), 101174. https://doi.org/10.1016/j.redox.2019.101174 doi: 10.1016/j.redox.2019.101174
|
| [21] |
L. Halim, M. Romano, R. McGregor, I. Correa, P. Pavlidis, N. Grageda, et al., An atlas of human regulatory T helper-like cells reveals features of Th2-like tregs that support a tumorigenic environment, Cell Rep., 20 (2017), 757–770. https://doi.org/10.1016/j.celrep.2017.06.079 doi: 10.1016/j.celrep.2017.06.079
|
| [22] |
L. Le, Y. Tokumaru, M. Oshi, M. Asaoka, L. Yan, I. Endo, et al., Th2 cell infiltrations predict neoadjuvant chemotherapy response of estrogen receptor-positive breast cancer, Gland Surg., 10 (2021), 154–165. https://doi.org/10.21037/gs-20-571 doi: 10.21037/gs-20-571
|
| [23] |
S. Schreiber, C. M. Hammers, A. J. Kaasch, B. Schraven, A. Dudeck, S. Kahlfuss, Metabolic interdependency of Th2 cell-mediated type 2 immunity and the tumor microenvironment, Front. Immunol., 12 (2021), 632581. https://doi.org/10.3389/fimmu.2021.632581 doi: 10.3389/fimmu.2021.632581
|
| [24] |
B. Marotti, A. Rocco, P. De. Colibus, D. Compare, G. de. Nucci, S. Staibano, et al., Interleukin-13 mucosal production in Helicobacter pylori-related gastric diseases, Dig. Liver Dis., 40 (2008), 240–247. https://doi.org/10.1016/j.dld.2007.11.02 doi: 10.1016/j.dld.2007.11.02
|
Figures(7) / Tables(2)

Yuan Yang, Yuwei Ye, Min Liu, Ya Zheng, Guozhi Wu, Zhaofeng Chen, Yuping Wang, Qinghong Guo, Rui Ji, Yongning Zhou. Family with sequence similarity 153 member B as a potential prognostic biomarker of gastric cancer[J]. Mathematical Biosciences and Engineering, 2022, 19(12): 12581-12600. doi: 10.3934/mbe.2022587







 DownLoad:
DownLoad: