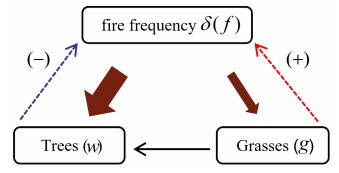
In this paper, we study the general mechanism of Turing-pattern in a tree-grass competition model with cross diffusion and time delay. The properties of four equilibrium points, the existence of Hopf bifurcation and the sufficient conditions for Turing instability caused by cross-diffusion are analyzed, respectively. The amplitude equation of tree-grass competition model is derived by using multi-scale analysis method, and its nonlinear stability is studied. The sensitivity analysis also verified that fire frequency plays a key role in tree-grass coexistence equilibrium. Finally, the Turing pattern of tree-grass model obtained by numerical simulation is consistent with the spatial structure of tree-grass density distribution observed in Hulunbuir grassland, China.
Citation: Rina Su, Chunrui Zhang. The generation mechanism of Turing-pattern in a Tree-grass competition model with cross diffusion and time delay[J]. Mathematical Biosciences and Engineering, 2022, 19(12): 12073-12103. doi: 10.3934/mbe.2022562
In this paper, we study the general mechanism of Turing-pattern in a tree-grass competition model with cross diffusion and time delay. The properties of four equilibrium points, the existence of Hopf bifurcation and the sufficient conditions for Turing instability caused by cross-diffusion are analyzed, respectively. The amplitude equation of tree-grass competition model is derived by using multi-scale analysis method, and its nonlinear stability is studied. The sensitivity analysis also verified that fire frequency plays a key role in tree-grass coexistence equilibrium. Finally, the Turing pattern of tree-grass model obtained by numerical simulation is consistent with the spatial structure of tree-grass density distribution observed in Hulunbuir grassland, China.

| [1] |
J. I. House, S. Archer, D. D. Breshears, R. J. Scholes, Conundrums in mixed woody-herbaceous plant systems, J. Biogeogr., 30 (2003), 1763–1777. https://doi.org/10.1046/j.1365-2699.2003.00873.x doi: 10.1046/j.1365-2699.2003.00873.x

|
| [2] |
M. Sankaran, J. Ratnam, N. P. Hanan, Tree-grass co-existence in savannas revisited-insights from an examination of assumptions and mechanisms invoked in existing models, Ecol. Lett., 7 (2004), 480–490. https://doi.org/10.1111/j.1461-0248.2004.00596.x doi: 10.1111/j.1461-0248.2004.00596.x
|
| [3] |
F. S. Gilliam, W. J. Platt, R. K. Peet, Natural disturbances and the physiognomy of pine savannas: a phenomenological model, Appl. Veg. Sci., 9 (2006), 83–96. https://doi.org/10.1111/j.1654-109X.2006.tb00658.x doi: 10.1111/j.1654-109X.2006.tb00658.x
|
| [4] |
P. D'Odorico, L. Francesco, L. Ridolfi, A probabilistic analysis of fire-induced tree-grass coexistence in savannas, Am. Nat., 167 (2006), E79–E87. https://doi.org/10.1086/500617 doi: 10.1086/500617
|
| [5] |
W. J. Bond, F. Woodward, G. Midgley, The global distribution of ecosystems in a world without fire, New Phytol., 165 (2005), 525–538. https://doi.org/10.1111/j.1469-8137.2004.01252.x doi: 10.1111/j.1469-8137.2004.01252.x
|
| [6] |
B. Beckage, L. J. Gross, Grass feedbacks on fire stabilize Savannas, Ecol. Modell., 222 (2011), 2227–2233. https://doi.org/10.1016/j.ecolmodel.2011.01.015 doi: 10.1016/j.ecolmodel.2011.01.015
|
| [7] | A. M. Gill, Adaptive responses of Australian vascular plant species to fires, Aust. Acad. Sci., (1981), 243–272. |
| [8] |
R. E. Keane, K. C. Ryan, T. T. Veblen, Cascading effects of fire exclusion in rocky mountain ecosystems: a literature review, USDA Forest Serv. General Tech. Rep., 91 (2002), 1–31. https://doi.org/10.2737/RMRS-GTR-91 doi: 10.2737/RMRS-GTR-91
|
| [9] | R. A. Bradstock, M. A. Gill, R. J. Williams, Flammable Australia: their roles in understanding and predicting biotic responses to fire regimes from individuals to landscapes, CSIRO Publishing, Colingwood Australia, 2012. https://doi.org/10.1007/978-1-4612-0873-0 |
| [10] |
W. A. Hoffmann, Fire and population dynamics of woody plants in a neotropical savanna: matrix model projections, Ecology, 80 (1999), 1354–1369. https://doi.org/10.2307/177080 doi: 10.2307/177080
|
| [11] |
B. Beckage, L. J. Gross, W. J. Platt, Modelling responses of pine Savannas to climate change and large-scale disturbance, Appl. Veg. Sci., 9 (2006), 75–82. https://doi.org/10.1111/j.1654-109X.2006.tb00657.x doi: 10.1111/j.1654-109X.2006.tb00657.x
|
| [12] |
W. Beckage, W. Platt, L. J. Gross, Vegetation, fire, and feedbacks: a disturbance-mediated model of Savannas, Am. Nat., 174 (2009), 805–818. https://doi.org/10.2307/27735896 doi: 10.2307/27735896
|
| [13] | T. D. Johan, Toit, R. Kevin, C. B. Harry, The kruger experience: ecology and management of savanna heterogeneity, Island Press, Washington, 2003. |
| [14] | M. Mermoz, T. Kitzberger, T. T. Veblen, Landscape influences on occurrence and spread of wildfires in patagonian forests and shrublands, Ecology, 86 (2005), 2705–2715. |
| [15] |
J. B. William, What limits trees in $C_4$ grasslands and Savannas, Annu. Rev. Ecol. Evol. Syst., 39 (2008), 641–659. https://doi.org/10.1146/annurev.ecolsys.39.110707.173411 doi: 10.1146/annurev.ecolsys.39.110707.173411
|
| [16] |
E. R. Caroline, A. T. Michael, M. Sankaran, S. I. Higgins, Savanna vegetation-fire-climate relationships differ among continents, Science, 343 (2014), 548–552. https://doi.org/10.1126/science.1247355 doi: 10.1126/science.1247355
|
| [17] | Q. Ouyang, Nonlinear Science and the Pattern Dynamics Introduction, Peking University Press, Beijing, 2010. |
| [18] |
C. Zhang, B. Zheng, R. Su, Realizability of the normal forms for the non-semisimple $1:1$ resonant Hopf bifurcation in a vector field, Commun. Nonlinear Sci. Numer. Simul., 91 (2020), 105407. https://doi.org/10.1016/j.cnsns.2020.105407 doi: 10.1016/j.cnsns.2020.105407
|
| [19] |
C. Zhang, B. Zheng, Steady state bifurcation and patterns of reaction-diffusion equations, Int. J. Bifurcat. Chaos, 30 (2020), 2050215. https://doi.org/10.1142/S0218127420502156 doi: 10.1142/S0218127420502156
|
| [20] |
W. Jiang, Q. An, J. Shi, Formulation of the normal form of Turing-Hopf bifurcation in partial functional differential equations, J. Differ. Equ., 268 (2020), 6067–6102. https://doi.org/10.1016/j.jde.2019.11.039 doi: 10.1016/j.jde.2019.11.039
|
| [21] |
K. Yun, K. Sourav, Sasmal, M. Komi, A two-patch prey-predator model with predator dispersal driven by the predation strength, Math. Biosci. Eng., 14 (2017), 843–880. https://doi.org/10.3934/mbe.2017046 doi: 10.3934/mbe.2017046
|
| [22] |
K. Kim, W. Choi, Local dynamics and coexistence of predator-prey model with directional dispersal of predator, Math. Biosci. Eng., 17 (2020), 6737–6755. https://doi.org/10.3934/mbe.2020351 doi: 10.3934/mbe.2020351
|
| [23] |
Q. Din, M. S. Shabbir, M. A. Khan, K. Ahmad, Bifurcation analysis and chaos control for a plant-herbivore model with weak predator functional response, J. Biol. Dyn., 13 (2019), 481–501. https://doi.org/10.1080/17513758.2019.1638976 doi: 10.1080/17513758.2019.1638976
|
| [24] |
A. M. Mahmoud, I. A. Ismail, A. A. Farah, H. Mohd, Codimension one and two bifurcations of a discrete-time fractional-order SEIR measles epidemic model with constant vaccination, Chaos Soliton. Fract., 140 (2020), 110104. https://doi.org/10.1016/j.chaos.2020.110104 doi: 10.1016/j.chaos.2020.110104
|
| [25] |
H. Huo, P. Yang, H. Xiang, Dynamics for an SIRS epidemic model with infection age and relapse on a scale-free network, J. Franklin Inst., 356 (2019), 7411–7443. https://doi.org/10.1016/j.jfranklin.2019.03.034 doi: 10.1016/j.jfranklin.2019.03.034
|
| [26] |
X. Yang, F. Liu, Q. Wang, H. Wang, Dynamics analysis for a discrete dynamic competition model, Adv. Differ. Equ., 2019 (2019), 1–17. https://doi.org/10.1186/s13662-019-2149-6 doi: 10.1186/s13662-019-2149-6
|
| [27] |
H. Kato, T. Takada, Stability and bifurcation analysis of a ratio-dependent community dynamics model on Batesian mimicry, J. Math. Biol., 79 (2019), 329–368. https://doi.org/10.1007/s00285-019-01359-y doi: 10.1007/s00285-019-01359-y
|
| [28] |
Y. Song, Hopf bifurcation and spatio-temporal patterns indelay-coupled van der Pol oscillators, Nonlinear Dyn., 63 (2011), 223–237. https://doi.org/10.1007/s11071-010-9799-y doi: 10.1007/s11071-010-9799-y
|
| [29] |
J. Lin, R. Xu, L. Li, Turing-Hopf bifurcation of reaction-diffusion neural networks with leakage delay, Commun. Nonlinear Sci. Numer. Simul., 85 (2020), 105241. https://doi.org/10.1016/j.cnsns.2020.105241 doi: 10.1016/j.cnsns.2020.105241
|
| [30] |
R. J. Scholes, S. R. Archer, Tree-grass interactions in Savannas, Ann. Rev. Ecol. Syst., 28 (1997), 517–544. https://doi.org/10.1146/annurev.ecolsys.28.1.517 doi: 10.1146/annurev.ecolsys.28.1.517
|
| [31] |
S. I. Higgins, W. J. Bond, W. Trollope, Fire, resprouting and variability: a recipe for grass-tree coexistence in Savanna, J. Ecol., 88 (2000), 213–229. https://doi.org/10.1046/j.1365-2745.2000.00435.x doi: 10.1046/j.1365-2745.2000.00435.x
|
| [32] |
J. Shi, Z. Xie, K. Little, Cross-diffusion induced instability and stability in reaction-diffusion systems, J. Appl. Anal. Comput., 1 (2011), 95–119. https://doi.org/10.2337/diab.31.7.585 doi: 10.2337/diab.31.7.585
|
| [33] |
Q. Li, Z. Liu, S. Yuan, Cross-diffusion induced Turing instability for a competition model with saturation effect, J. Math. Comput., 347 (2019), 64–77. https://doi.org/10.1016/j.amc.2018.10.071 doi: 10.1016/j.amc.2018.10.071
|
| [34] |
S. Chen, J. Shi, J. Wei, Time delay-induced instabilities and Hopf bifurcations in general reaction–diffusion systems, J. Nonlinear Sci., 23 (2013), 1–38. https://doi.org/10.1007/s00332-012-9138-1 doi: 10.1007/s00332-012-9138-1
|
| [35] |
V. Yatat, P. Couteron, J. J. Tewa, An impulsive modelling framework of fire occurrence in a size structured model of tree-grass interactions for savanna ecosystems, J. Math. Biol., 74 (2017), 1425–1482. https://doi.org/10.1007/s00285-016-1060-y doi: 10.1007/s00285-016-1060-y
|
| [36] |
Y. Su, J. Wei, J. Shi, Hopf bifurcations in a reaction-diffusion population model with delay effect, J. Differ. Equ., 247 (2009), 1156–1184. https://doi.org/10.1016/j.jde.2009.04.017 doi: 10.1016/j.jde.2009.04.017
|
| [37] |
F. Wei, Q. Fu, Hopf bifurcation and stability for predator-prey systems with Beddington-DeAngelis type functional response and stage structure for prey incorporating refuge, Appl. Math. Model., 40 (2016), 126–134. https://doi.org/10.1016/j.apm.2015.04.042 doi: 10.1016/j.apm.2015.04.042
|
| [38] |
F. Accatino, C. De Michele, R. Vezzoli, D. Donzelli, R. J. Scholes, Tree-grass co-existence in Savanna: interactions of rain and fire, J. Theor. Biol., 267 (2010), 235–242. https://doi.org/10.1016/j.jtbi.2010.08.012 doi: 10.1016/j.jtbi.2010.08.012
|
| [39] |
M. Garvie, Finite difference schemes for reaction-diffusion equations modeling predator-prey interactions in MATLAB, Bull. Math. Biol., 69 (2007), 931–956. https://doi.org/10.1007/s11538-006-9062-3 doi: 10.1007/s11538-006-9062-3
|
















Figures(17) / Tables(2)

Rina Su, Chunrui Zhang. The generation mechanism of Turing-pattern in a Tree-grass competition model with cross diffusion and time delay[J]. Mathematical Biosciences and Engineering, 2022, 19(12): 12073-12103. doi: 10.3934/mbe.2022562







 DownLoad:
DownLoad: