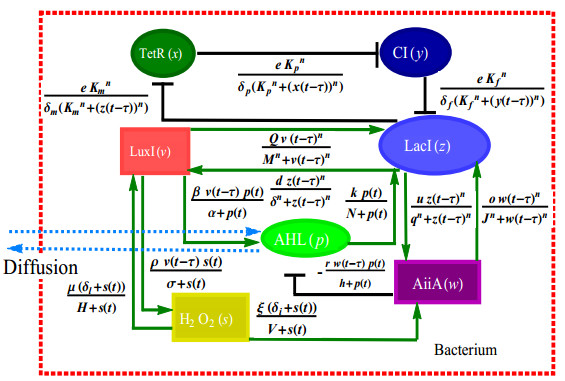
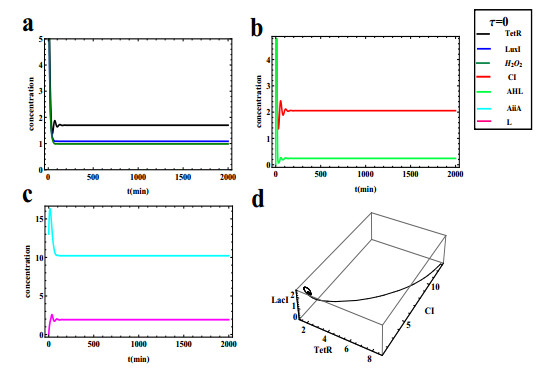
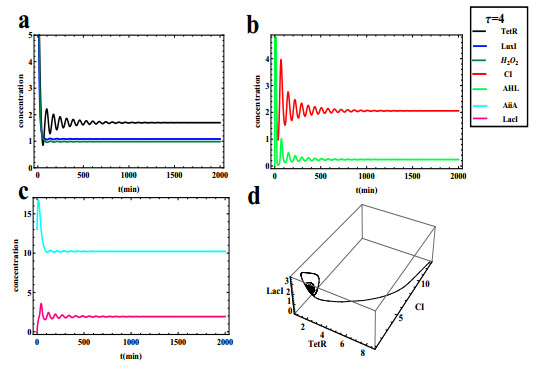
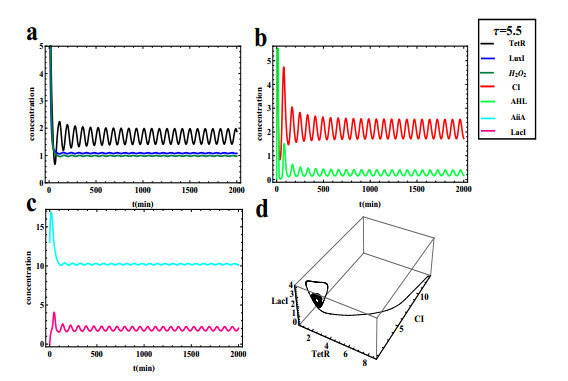
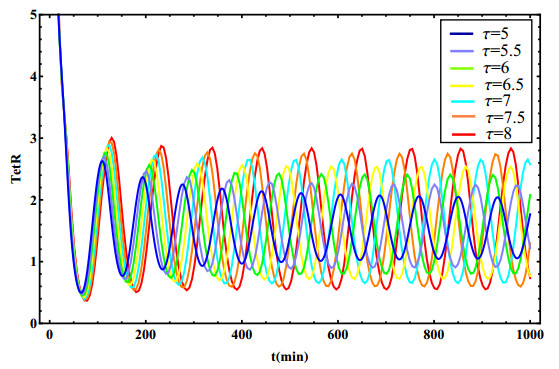
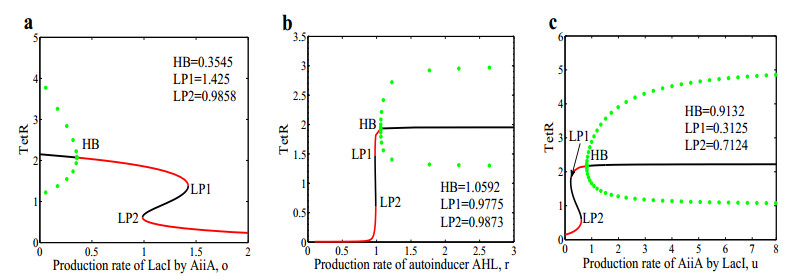
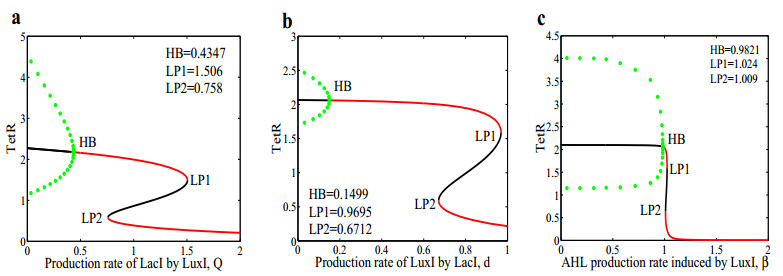
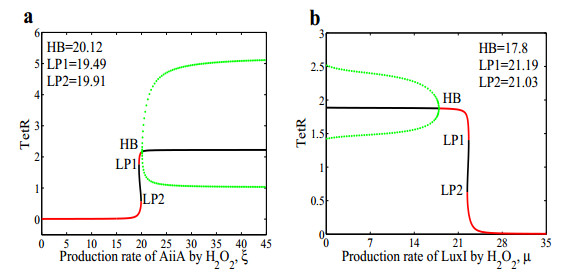
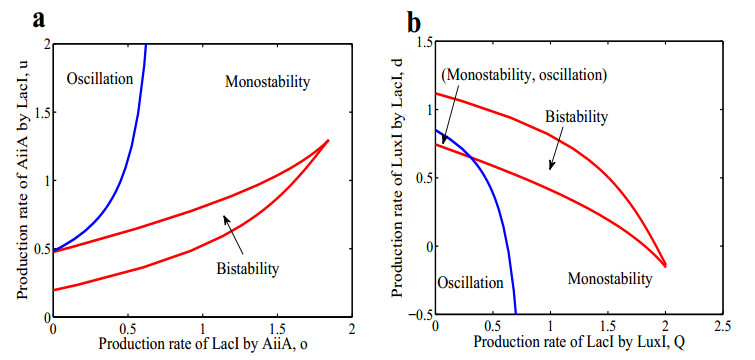
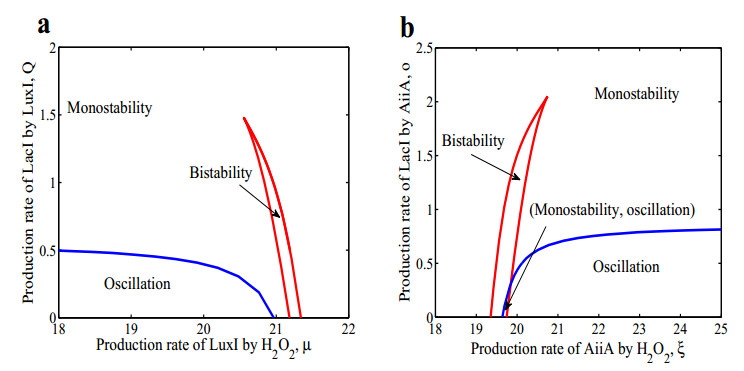
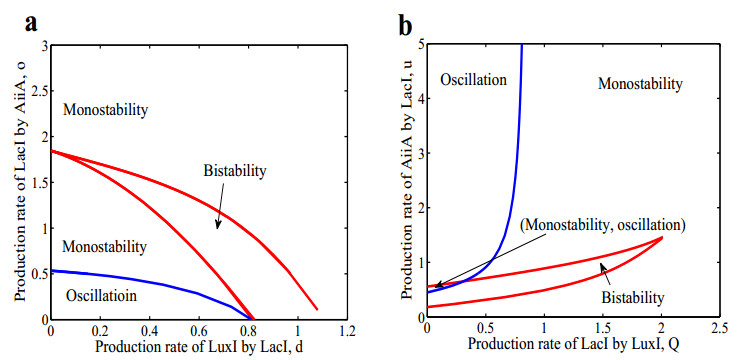
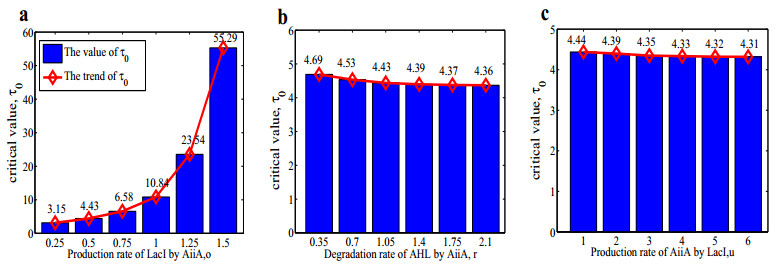
The dynamical behaviors of the quorum sensing (QS) system are closely related to the release drugs and control the PH value in microorganisms and plants. However, the effect of the main molecules AiiA, LuxI, H$ _2 $O$ _2 $, and time delayed individual and combinatorial perturbation on the QS system dynamics and the above-mentioned biological phenomena is still unclear, which are seen as a key consideration in our paper. This paper formulates a QS computational model by incorporating these several substances. First, for the protein production time delay, a critical value is given by Hopf bifurcation theory. It is found that a larger time delay can lead to a larger amplitude and a longer period. This indicates that the length of time for protein synthesis has a regulatory effect on the release of drugs from the bacterial population. Second, hen the concentrations of AiiA, LuxI, and H$ _2 $O$ _2 $ is modulated individually, the QS system undergoes periodic oscillation and bistable state. Meanwhile, oscillatory and bistable regions can be significantly affected by simultaneously perturbing any two parameters related to AiiA, LuxI, and H$ _2 $O$ _2 $. This means that the individual or simultaneous changes of the three intrinsic molecular concentrations can effectively control the drugs release and the PH value in microorganisms and plants. Finally, the sensitivity relationship between the critical value of the delay and AiiA, LuxI, H$ _2 $O$ _2 $ parameters is analyzed.
Citation: Menghan Chen, Haihong Liu, Ruiqi Wang. Dynamical behaviors of quorum sensing network mediated by combinatorial perturbation[J]. Mathematical Biosciences and Engineering, 2022, 19(5): 4812-4840. doi: 10.3934/mbe.2022225
The dynamical behaviors of the quorum sensing (QS) system are closely related to the release drugs and control the PH value in microorganisms and plants. However, the effect of the main molecules AiiA, LuxI, H$ _2 $O$ _2 $, and time delayed individual and combinatorial perturbation on the QS system dynamics and the above-mentioned biological phenomena is still unclear, which are seen as a key consideration in our paper. This paper formulates a QS computational model by incorporating these several substances. First, for the protein production time delay, a critical value is given by Hopf bifurcation theory. It is found that a larger time delay can lead to a larger amplitude and a longer period. This indicates that the length of time for protein synthesis has a regulatory effect on the release of drugs from the bacterial population. Second, hen the concentrations of AiiA, LuxI, and H$ _2 $O$ _2 $ is modulated individually, the QS system undergoes periodic oscillation and bistable state. Meanwhile, oscillatory and bistable regions can be significantly affected by simultaneously perturbing any two parameters related to AiiA, LuxI, and H$ _2 $O$ _2 $. This means that the individual or simultaneous changes of the three intrinsic molecular concentrations can effectively control the drugs release and the PH value in microorganisms and plants. Finally, the sensitivity relationship between the critical value of the delay and AiiA, LuxI, H$ _2 $O$ _2 $ parameters is analyzed.

| [1] |
S. Ahmed, M. Rudden, T. J. Smyth, J. S. Dooley, R. Marchant, I. M. Banat, Natural quorum sensing inhibitors effectively downregulate gene expression of pseudomonas aeruginosa virulence factors, Appl. Microbiol. Biotechnol., 103 (2019), 3521–3535. https://doi.org/10.1007/s00253-019-09618-0 doi: 10.1007/s00253-019-09618-0

|
| [2] |
H. Wang, W. Chu, C. Ye, G. Bruno, H. Tao, M. Wang, et al., Chlorogenic acid attenuates virulence factors and pathogenicity of pseudomonas aeruginosa by regulating quorum sensing, Appl. Microbiol. Biotechnol., 103 (2019), 903–915. https://doi.org/10.1007/s00253-018-9482-7 doi: 10.1007/s00253-018-9482-7
|
| [3] |
A. Gupta, I. M. B. Reizman, C. R. Reisch, K. L. Prather, Dynamic regulation of metabolic flux in engineered bacteria using a pathway-independent quorum-sensing circuit, Nat. Biotechnol., 35 (2017), 273. https://doi.org/10.1038/nbt.3796 doi: 10.1038/nbt.3796
|
| [4] |
H. Morten, W. Hong, A. Jens Bo, R. Kathrin, T. B. Rasmussen, B. Niels, et al., Attenuation of pseudomonas aeruginosa virulence by quorum sensing inhibitors, EMBO J., 22 (2014), 3803–3815. https://doi.org/10.1093/emboj/cdg366 doi: 10.1093/emboj/cdg366
|
| [5] |
D. Viducic, K. Murakami, T. Amoh, T. Ono, Y. Miyake, Role of the interplay between quorum sensing regulator vqsr and the pseudomonas quinolone signal in mediating carbapenem tolerance in pseudomonas aeruginosa, Res. Microbiol., 168 (2017), 450–460. https://doi.org/10.1016/j.resmic.2017.02.007 doi: 10.1016/j.resmic.2017.02.007
|
| [6] |
K. C. Tu, B. L. Bassler, Multiple small rnas act additively to integrate sensory information and control quorum sensing in vibrio harveyi, Genes Dev., 21 (2007), 221–33. https://doi.org/10.1101/gad.1502407 doi: 10.1101/gad.1502407
|
| [7] |
Q. Zhao, C. Zhang, Z. Jia, Y. Huang, H. Li, S. Song, Involvement of calmodulin in regulation of primary root elongation by n-3-oxo-hexanoyl homoserine lactone in arabidopsis thaliana, Front. Plant Sci., 5 (2016), 807. https://doi.org/10.3389/fpls.2014.00807 doi: 10.3389/fpls.2014.00807
|
| [8] |
W. L. Ng, B. L. Bassler, Bacterial quorum-sensing network architectures, Annu. Rev. Genet., 43 (2009), 197. https://doi.org/10.1146/annurev-genet-102108-134304 doi: 10.1146/annurev-genet-102108-134304
|
| [9] |
C. Fuqua, M. R. Parsek, E. P. Greenberg, Regulation of gene expression by cell-to-cell communication: acyl-homoserine lactone quorum sensing–-annual review of genetics, Annu. Rev. Genet., 35 (2001), 439–468. https://doi.org/10.1146/annurev.genet.35.102401.090913 doi: 10.1146/annurev.genet.35.102401.090913
|
| [10] |
S. Zhou, Synthetic biology: bacteria synchronized for drug delivery, Nature, 536 (2016), 33–34. https://doi.org/10.1038/nature18915 doi: 10.1038/nature18915
|
| [11] |
S. Majumdar, S. Mondal, Conversation game: talking bacteria, Cell Commun. Signal, 10 (2016), 331–335. https://doi.org/10.1007/s12079-016-0333-y doi: 10.1007/s12079-016-0333-y
|
| [12] |
S. Majumdar, S. Pal, Quorum sensing: a quantum perspective, Cell Commun. Signal, 10 (2016), 173–175. https://doi.org/10.1007/s12079-016-0348-4 doi: 10.1007/s12079-016-0348-4
|
| [13] |
B. Novák, J. J. Tyson, Design principles of biochemical oscillators, Nat. Rev. Mol. Cell Biol., 9 (2008), 981–991. https://doi.org/10.1038/nrm2530 doi: 10.1038/nrm2530
|
| [14] |
J. Sneyd, J. M. Han, L. Wang, J. Chen, X. Yang, A. Tanimura, et al., On the dynamical structure of calcium oscillations, Proc. Natl. Acad. Sci. USA, 114 (2017), 1456–1461. https://doi.org/10.1073/pnas.1614613114 doi: 10.1073/pnas.1614613114
|
| [15] |
M. Chen, J. Ji, H. Liu, F. Yan, Periodic oscillations in the quorum-sensing system with time delay, Int. J. Bifurcation Chaos, 30 (2020), 2050127. https://doi.org/10.1142/S0218127420501278 doi: 10.1142/S0218127420501278
|
| [16] |
M. Chen, H. Liu, F. Yan, Modeling and analyzing biological oscillations in quorum sensing networks, IET Syst. Biol., 14 (2020), 190–199. https://doi.org/10.1049/iet-syb.2019.0079 doi: 10.1049/iet-syb.2019.0079
|
| [17] |
T. Bánsági Jr, A. F. Taylor, Switches induced by quorum sensing in a model of enzyme-loaded microparticles, J. R. Soc. Interface, 15 (2018), 20170945. https://doi.org/10.1098/rsif.2017.0945 doi: 10.1098/rsif.2017.0945
|
| [18] |
D. Ran, J. Mao, C. Zhan, C. Xie, H. Ruan, M. Ying, et al., D-retroenantiomer of quorum-sensing peptide-modified polymeric micelles for brain tumor-targeted drug delivery, ACS Appl. Mater. Interfaces, 9 (2017), 25672–25682. https://doi.org/10.1021/acsami.7b03518 doi: 10.1021/acsami.7b03518
|
| [19] |
D. Gordon, Bacteria as drug delivery systems, Gastroenterology, 119 (2000), 1187–1188. https://doi.org/10.1016/S0016-5085(00)70288-5 doi: 10.1016/S0016-5085(00)70288-5
|
| [20] |
M. O. Din, T. Danino, A. Prindle, M. Skalak, J. Selimkhanov, K. Allen, et al., Synchronized cycles of bacterial lysis for in vivo delivery, Nature, 536 (2016), 81–85. https://doi.org/10.1038/nature18930 doi: 10.1038/nature18930
|
| [21] |
M. B. Elowitz, S. Leibler, A synthetic oscillatory network of transcriptional regulators, Nature, 403 (2000), 335–338. https://doi.org/10.1038/35002125 doi: 10.1038/35002125
|
| [22] |
J. Garcia-Ojalvo, M. B. Elowitz, S. H. Strogatz, Modeling a synthetic multicellular clock: repressilators coupled by quorum sensing, Proc. Natl. Acad. Sci. USA., 101 (2004), 10955–10960. https://doi.org/10.1073/pnas.0307095101 doi: 10.1073/pnas.0307095101
|
| [23] |
I. Potapov, E. Volkov, A. Kuznetsov, Dynamics of coupled repressilators: the role of mrna kinetics and transcription cooperativity, Phys. Rev. E, 83 (2011), 031901. https://doi.org/10.1103/PhysRevE.83.031901 doi: 10.1103/PhysRevE.83.031901
|
| [24] |
I. Potapov, B. Zhurov, E. Volkov, "Quorum sensing" generated multistability and chaos in a synthetic genetic oscillator, Chaos, 22 (2012), 023117. https://doi.org/10.1063/1.4705085 doi: 10.1063/1.4705085
|
| [25] |
V. Agrawal, S. S. Kang, S. Sinha, Realization of morphing logic gates in a repressilator with quorum sensing feedback, Phys. Lett. A, 378 (2014), 1099–1103. https://doi.org/10.1016/j.physleta.2014.02.015 doi: 10.1016/j.physleta.2014.02.015
|
| [26] |
T. Gedeon, M. Pernarowski, A. Wilander, Cyclic feedback systems with quorum sensing coupling, Bull. Math. Biol., 78 (2016), 1291–1317. https://doi.org/10.1007/s11538-016-0187-8 doi: 10.1007/s11538-016-0187-8
|
| [27] |
L. Guo, M. Hu, Z. Xu, A. Hu, Synchronization and chaos control by quorum sensing mechanism, Nonlinear Dyn., 73 (2013), 1253–1269. https://doi.org/10.1007/s11071-013-0769-z doi: 10.1007/s11071-013-0769-z
|
| [28] |
Ž. Pušnik, M. Mraz, N. Zimic, M. Moškon, Computational analysis of viable parameter regions in models of synthetic biological systems, J. Biol. Eng., 13 (2019), 1–21. https://doi.org/10.1186/s13036-019-0205-0 doi: 10.1186/s13036-019-0205-0
|
| [29] |
R. L. Ulrich, D. DeShazer, E. E. Brueggemann, H. B. Hines, P. C. Oyston, J. A. Jeddeloh, Role of quorum sensing in the pathogenicity of burkholderia pseudomallei, J. Med. Microbiol., 53 (2004), 1053–1064. https://doi.org/10.1099/jmm.0.45661-0 doi: 10.1099/jmm.0.45661-0
|
| [30] | S. J. Park, S. Y. Park, C. M. Ryu, S. H. Park, J. K. Lee, The role of aiia, a quorum-quenching enzyme from bacillus thuringiensis, on the rhizosphere competence, J. Microbiol. Biotechnol., 18 (2008), 1518–1521. |
| [31] |
J. Wang, J. Zhang, Z. Yuan, T. Zhou, Noise-induced switches in network systems of the genetic toggle switch, BMC Syst. Biol., 1 (2007), 1–14. https://doi.org/10.1186/1752-0509-1-50 doi: 10.1186/1752-0509-1-50
|
| [32] |
A. Prindle, J. Selimkhanov, H. Li, I. Razinkov, L. S. Tsimring, J. Hasty, Rapid and tunable post-translational coupling of genetic circuits, Nature, 508 (2014), 387–391. https://doi.org/10.1038/nature13238 doi: 10.1038/nature13238
|
| [33] |
N. A. Monk, Oscillatory expression of hes1, p53, and nf-b driven by transcriptional time delays, Curr. Biol., 13 (2003), 1409–1413. https://doi.org/10.1016/S0960-9822(03)00494-9 doi: 10.1016/S0960-9822(03)00494-9
|
| [34] |
Y. Zhang, H. Liu, F. Yan, J. Zhou, Oscillatory dynamics of p38 activity with transcriptional and translational time delays, Sci. Rep., 7 (2017), 11495. https://doi.org/10.1038/s41598-017-11149-5 doi: 10.1038/s41598-017-11149-5
|
| [35] |
C. Wang, H. Liu, J. Zhou, Oscillatory dynamics of p53 genetic network induced by feedback loops and time delays, IEEE Trans. Nanobiosci., 18 (2019), 611–621. https://doi.org/10.1109/TNB.2019.2924079 doi: 10.1109/TNB.2019.2924079
|
| [36] |
E. Batchelor, C. S. Mock, I. Bhan, A. Loewer, G. Lahav, Recurrent initiation: A mechanism for triggering p53 pulses in response to dna damage, Mol. Cell, 30 (2008), 277–289. https://doi.org/10.1016/j.molcel.2008.03.016 doi: 10.1016/j.molcel.2008.03.016
|
| [37] |
T. Danino, O. Mondragnpalomino, L. Tsimring, J. Hasty, A synchronized quorum of genetic clocks, Nature, 463 (2010), 326–30. https://doi.org/10.1038/nature08753 doi: 10.1038/nature08753
|
| [38] | B. Borek, J. Hasty, L. Tsimring, Turing patterning using gene circuits with gas-induced degradation of quorum sensing molecules, PloS one, 11 (2016), e0153679. https://doi.org/10.1371/journal.pone.0160272 https://doi.org/10.1371/journal.pone.0160272 |
| [39] |
B. Huang, M. Lu, D. Jia, E. Ben-Jacob, H. Levine, J. N. Onuchic, Interrogating the topological robustness of gene regulatory circuits by randomization, PloS Comput. Biol., 13 (2017), e1005456. https://doi.org/10.1371/journal.pcbi.1005456 doi: 10.1371/journal.pcbi.1005456
|
| [40] |
C. Gao, J. Ji, F. Yan, H. Liu, Oscillation induced by Hopf bifurcation in the p53–Mdm2 feedback module, IET Syst. Biol., 13 (2019), 251–259. https://doi.org/10.1049/iet-syb.2018.5092 doi: 10.1049/iet-syb.2018.5092
|
| [41] |
S. Bakshi, H. Choi, J. Mondal, J. C. Weisshaar, Time-dependent effects of transcription-and translation-halting drugs on the spatial distributions of the e scherichia coli chromosome and ribosomes, Mol. Microbiol., 94 (2014), 871–887. https://doi.org/10.1111/mmi.12805 doi: 10.1111/mmi.12805
|
| [42] |
E. A. Campbell, N. Korzheva, A. Mustaev, K. Murakami, S. Nair, A. Goldfarb, et al., Structural mechanism for rifampicin inhibition of bacterial RNA polymerase, Cell, 104 (2001), 901–912. https://doi.org/10.1016/S0092-8674(01)00286-0 doi: 10.1016/S0092-8674(01)00286-0
|
| [43] |
F. Schluenzen, C. Takemoto, D. N. Wilson, T. Kaminishi, J. M. Harms, K. Hanawasuetsugu, et al., The antibiotic kasugamycin mimics mrna nucleotides to destabilize tRNA binding and inhibit canonical translation initiation, Nat. Struct. Mol. Biol., 13 (2006), 871. https://doi.org/10.1038/nsmb1145 https://doi.org/10.1038/nsmb1145 doi: 10.1038/nsmb1145
|
| [44] |
J. L. Hansen, P. B. Moore, T. A. Steitz, Structures of five antibiotics bound at the peptidyl transferase center of the large ribosomal subunit, J. Mol. Biol., 330 (2003), 1061–1075. https://doi.org/10.1142/11705 doi: 10.1142/11705
|
| [45] |
L. Ouyang, L. Li, Effects of an inducible aiia gene on disease resistance in eucalyptus urophylla$\times$ eucalyptus grandis, Transgenic. Res., 25 (2016), 441–452. https://doi.org/10.1007/s11248-016-9940-x doi: 10.1007/s11248-016-9940-x
|
| [46] |
J. Pan, T. Huang, F. Yao, Z. Huang, C. A. Powell, S. Qiu, et al., Expression and characterization of aiia gene from bacillus subtilis bs-1, Microbiol. Res., 163 (2008), 711–716. https://doi.org/10.1016/j.micres.2007.12.002 doi: 10.1016/j.micres.2007.12.002
|
| [47] | F. Modarresi, O. Azizi, M. R. Shakibaie, M. Motamedifar, S. Mansouri, Cloning and expression of quorum sensing n-acyl-homoserine synthase (luxi) gene detected in acinetobacter baumannii, Iran. J. Microbiol., 8 (2016), 139. |
| [48] |
A. Prindle, P. Samayoa, I. Razinkov, T. Danino, L. S. Tsimring, J. Hasty, A sensing array of radically coupled genetic 'biopixels', Nature, 481 (2012), 39–44. https://doi.org/10.1038/nature10722 doi: 10.1038/nature10722
|
| [49] |
C. Li, L. Liu, T. Zhang, F. Yan, Hopf bifurcation analysis of a gene regulatory network mediated by small noncoding RNA with time delays and diffusion, Int. J. Bifurcation Chaos, 27 (2017), 1750194. https://doi.org/10.1142/S0218127417501942 doi: 10.1142/S0218127417501942
|
| [50] |
J. Müller, C. Kuttler, B. Hense, M. Rothballer, A. Hartmann, Cell–cell communication by quorum sensing and dimension-reduction Journal of mathematical biology, J. Math. Biol., 53 (2006), 672–702. https://doi.org/10.1007/s00285-006-0024-z doi: 10.1007/s00285-006-0024-z
|
| [51] |
B. Hense, C. Kuttler, J. Müller, M. Rothballer, A. Hartmann, J. U. Kreft, Does efficiency sensing unify diffusion and quorum sensing?, Nature Rev. Microbiol., 5 (2007), 230–239. https://doi.org/10.1038/nrmicro1600 doi: 10.1038/nrmicro1600
|
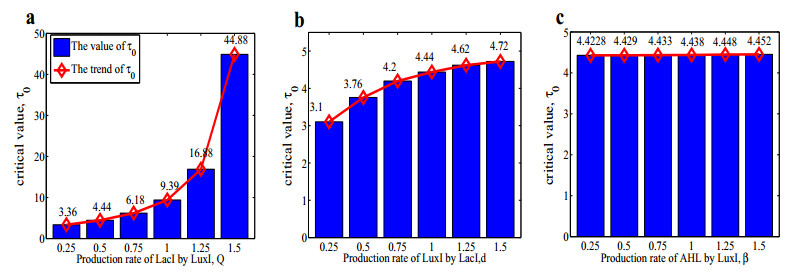
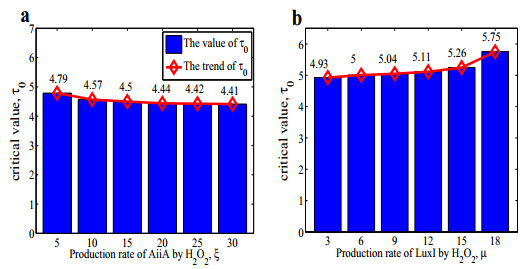
Figures(14) / Tables(1)

Menghan Chen, Haihong Liu, Ruiqi Wang. Dynamical behaviors of quorum sensing network mediated by combinatorial perturbation[J]. Mathematical Biosciences and Engineering, 2022, 19(5): 4812-4840. doi: 10.3934/mbe.2022225







 DownLoad:
DownLoad: