Citation: Ahsan H. Khandoker, Haitham M. Al-Angari, Andreas Voss, Steffen Schulz, Yoshitaka Kimura. Quantification of maternal-fetal cardiac couplings in normal and abnormal pregnancies applying high resolution joint symbolic dynamics[J]. Mathematical Biosciences and Engineering, 2020, 17(1): 802-813. doi: 10.3934/mbe.2020042

| [1] | A. H. Khandoker, S. Schulz, H. M. Al-Angari, et al, Alterations in maternal-fetal heart rate coupling strength and directions in abnormal fetuses, Front. Physiol., 10 (2019), 482. |
| [2] | C. Monk, W. P. Fifer, M. M. Myers, et al, Maternal stress responses and anxiety during pregnancy: Effects on fetal heart rate, Dev. Psychobiol., 36 (2000), 67-77. |
| [3] | D. J. Bekedam, E. J. Mulder, R. J. Snijders, et al, The effects of maternal hyperoxia on fetal breathing movements, body movements and heart rate variation in growth retarded fetuses, Early Hum. Dev., 27 (1991), 223-232. |
| [4] | P. Van Leeuwen, K. M. Gustafson, D. Cysarz, et al, Aerobic exercise during pregnancy and presence of fetal-maternal heart rate synchronization, PLoS One, 9 (2014), e106036. |
| [5] | A. H. Khandoker, F. Marzbanrad, A. Voss, et al, Analysis of maternal-fetal heart rate coupling directions with partial directed coherence, Biomed. Signal. Proces., 3 (2016), 25-30. |
| [6] | M. Riedl, P. Van Leeuwen, A. Suhrbier, et al, Testing foetal-maternal heart rate synchronization via model-based analyses, Phil. Trans. Math. Phys. Eng. Sci., 367 (2009), 1407-1421. |
| [7] | P. Van Leeuwen, D. Geue, M. Thiel, et al, Influence of paced maternal breathing on fetal-maternal heart rate coordination, Proc. Natl. Acad. Sci. U S A, 106 (2009), 13661-13666. |
| [8] | P. Ivanov, Q. D. Ma, R. P. Bartsch, Maternal-fetal heartbeat phase synchronization, Proc. Natl. Acad. Sci. U S A, 106 (2009), 13641-13642. |
| [9] | A. Porta and L. Faes, Assessing causality in brain dynamics and cardiovascular control, Phil. Trans. Math. Phys. Eng. Sci., 371(2013), 20120517. |
| [10] | S. Schulz and A. Voss, Cardiovascular and cardiorespiratory coupling analysis-State of the art and future perspectives, in Cardiovascular Oscillations (ESGCO), 2014 8th Conference of the European Study Group, (2014), 25-26. |
| [11] | S. Schulz, F. C. Adochiei, I. R. Edu, et al, Cardiovascular and cardiorespiratory coupling analyses: A review, Phil. Trans. Math. Phys. Eng. Sci., 371(2013): 20120191. |
| [12] | M. Baumert, T. Walther, J. Hopfe, et al, Joint symbolic dynamic analysis of beat-to-beat interactions of heart rate and systolic blood pressure in normal pregnancy, Med. Biol. Eng. Comput., 40 (2002), 241-245. |
| [13] | L. Faes, A. Porta and G. Nollo, Mutual nonlinear prediction as a tool to evaluate coupling strength and directionality in bivariate time series: comparison among different strategies based on k nearest neighbors, Phys. Rev. E, Stat. nonlin. Soft Matter phys., 78 (2008): 026201. |
| [14] | A. Porta, G. Baselli, F. Lombardi, et al, Conditional entropy approach for the evaluation of the coupling strength, Biol. Cybern., 81 (1999), 119-129. |
| [15] | M. G. Rosenblum and A. S. Pikovsky, Detecting direction of coupling in interacting oscillators, Phys. Rev. E Stat. nonlin. Soft Matter phys., 64 (2001), 045202. |
| [16] | T. Milde, K. Schwab, M. Walther, et al, Time-variant partial directed coherence in analysis of the cardiovascular system, A methodological study, Physiol. Meas., 32 (2011), 1787-1805. |
| [17] | S. Schulz, J. Haueisen, K. J. Bar, et al, High-resolution joint symbolic analysis to enhance classification of the cardiorespiratory system in patients with schizophrenia and their relatives, Phil. Trans. Math. Phys. Eng. Sci., 373 (2015), 2034. |
| [18] | S. Schulz, N. Tupaika, S. Berger, et al, Cardiovascular coupling analysis with high-resolution joint symbolic dynamics in patients suffering from acute schizophrenia, Physiol. Meas., 34 (2013), 883-901. |
| [19] | S. Reulecke, S. Schulz and A. Voss, Autonomic regulation during quiet and active sleep states in very preterm neonates, Front. Physiol., 3 (2012), 61. |
| [20] | A. H. Khandoker, S. Schulz, Y. Kimura, et al, Investigating maternal-fetal heart rate coupling by high resolution joint symbolic dynamics, Comput. Cardiol., 41 (2014), 441-444. |
| [21] | M. Sato, Y. Kimura, S. Chida, et al, A novel extraction method of fetal electrocardiogram from the composite abdominal signal, IEEE Trans. Biomed. Eng., 54 (2007), 49-58. |
| [22] | J. Pan and W. J. Tompkins, A Real-Time Qrs Detection Algorithm, IEEE Trans. Biomed. Eng., 32 (1985), 230-236. |
| [23] | T. Ohta, K. Okamura, Y. Kimura, et al, Alteration in the low-frequency domain in power spectral analysis of fetal heart beat fluctuations, Fetal Diagn. Ther., 14 (1999), 92-97. |
| [24] | Types of Heart Block, 2012. Available from: www.nhlbi.nih.gov/health/health-topics/topics/hb/types. |
| [25] | S. H. Lim, V. Anantharaman, W. S. Teo, et al, Comparison of treatment of supraventricular tachycardia by Valsalva maneuver and carotid sinus massage, Ann. Emerg. Med., 31 (1998), 30-35. |

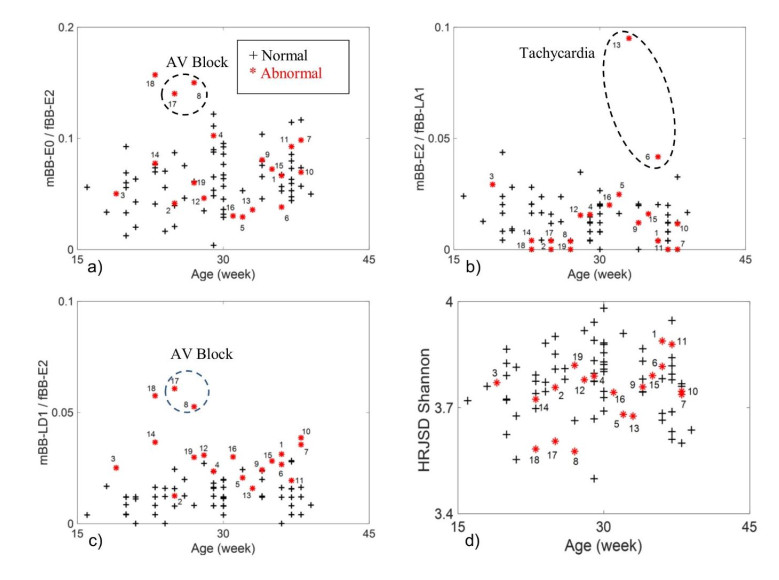
Figures(2) / Tables(6)

Ahsan H. Khandoker, Haitham M. Al-Angari, Andreas Voss, Steffen Schulz, Yoshitaka Kimura. Quantification of maternal-fetal cardiac couplings in normal and abnormal pregnancies applying high resolution joint symbolic dynamics[J]. Mathematical Biosciences and Engineering, 2020, 17(1): 802-813. doi: 10.3934/mbe.2020042







 DownLoad:
DownLoad: