Citation: Andrei Korobeinikov, Elena Shchepakina, Vladimir Sobolev. A black swan and canard cascades in an SIR infectious disease model[J]. Mathematical Biosciences and Engineering, 2020, 17(1): 725-736. doi: 10.3934/mbe.2020037

| [1] | E. Shchepakina and V. Sobolev, Black Swans and Canards in Laser and Combustion Models, in Singular Perturbation and Hysteresis (eds. M P Mortell et al.), SIAM, Philadelphia, (2005), 207-255. |
| [2] | E. Shchepakina, V. Sobolev and M. P. Mortell, Singular Perturbations: Introduction to System Order Reduction Methods with Applications, Springer Lecture Notes in Mathematics, Vol. 2114, Springer, Basel, 2014. |
| [3] | V. Sobolev, Geometry of Singular Perturbations: Critical Cases, Singular Perturbation and Hysteresis, (eds. M.P. Mortell et al.), SIAM, Philadelphia, (2005), 153-206. |
| [4] | N. N. Bogolyubov, On Some Statistical Methods in Mathematical Physics, Izd. Ukrain. Akad. Nauk, Kiev (1945). |
| [5] | N. N. Bogolyubov and Y. A. Mitropolsky, Asymptotic Methods in the Theory of Nonlinear Oscillations, Gordon and Breach, New York, 1961. |
| [6] | N. N. Bogolyubov and Y. A. Mitropolsky, The method of integral manifolds in nonlinear mechanics, Contributions to Differential Equations, 2, Wiley, New York, (1963), 123-196. |
| [7] | J. Hale, Integral manifolds of perturbed differential systems annals of mathematics, Ann. Math., 73 (1961), 496-531. |
| [8] | J. Hale, Oscillations in Nonlinear Systems, MacGraw-Hill, New York, 1963. |
| [9] | J. Hale, Ordinary Differential Equations, Wiley, New York, 1969. |
| [10] | K. V. Zadiraka, On the integral manifold of a system of differential equations containing a small parameter, Dokl. Akad. Nauk SSSR, 115 (1957), 646-649. |
| [11] | K. V. Zadiraka, On a non-local integral manifold of a singularly perturbed differential system, Ukrain. Math. Z., 17 (1965), 47-63; also AMS Transl. Ser. 2, 89 (1970), 29-49. |
| [12] | V. A. Sobolev and V. V. Strygin, Permissibility of changing over to precession equations of gyroscopic systems, Mech. Solids, 13 (1978), 10-17. |
| [13] | V. V. Strygin and V. A. Sobolev, Effect of geometric and kinetic parameters and energy dissipation on orientation stability of satellites with double spin, Cosmic Res., 14 (1976), 331-335. |
| [14] | V. V. Strygin and V. A. Sobolev, Asymptotic methods in the problem of stabilization of rotating bodies by using passive dampers, Mech. Solids, 5 (1977), 19-25. |
| [15] | N. Fenichel, Geometric singular perturbation theory for ordinary differential equations, J. Differ. Equations, 31 (1979), 53-98. |
| [16] |
C. K. R. T. Jones, Geometric singular perturbation theory, Lecture Notes in Mathematics, 1609 (1994), 44-118. doi: 10.1007/BFb0095239

|
| [17] | M. Krupa and P. Szmolyan, Extending geometric singular perturbation theory to nonhyperbolic points-fold and canard points in two dimensions, SIAM J. Appl. Math., 33 (2001), 286-314. |
| [18] | E. Shchepakina and V. Sobolev, Integral manifolds, canards and black swans, Nonlinear Anal. Theor. Methods Appl., 44 (2001), 897-908. |
| [19] | Z. Feng, Y. Yi and H. Zhu, Fast and slow dynamics of malaria and the s-gene frequency, J. Dyn. Differ. Equ., 16 (2004), 869-896. |
| [20] | M. Li, W. Liu, C. Shan, et al., Turning points and relaxation oscillation cycles in simple oscillation cycles in simple epidemic models, SIAM J. Appl. Math., 76 (2018), 663-687. |
| [21] | W. Liu, D. Xiao and Y. Yi, Relaxation oscillations in a class of predator-prey systems, J. Differ. Equations, 188 (2003), 306-331. |
| [22] | E. Benoit, J. L. Calot, F. Diener, et al., Chasse au canard, Collect. Math., 31-32(1-3) (1980-1981), 37-119. |
| [23] | E. Shchepakina, Black swans and canards in self-ignition problem, Nonlinear Anal-Real, 4 (2003), 45-50. |
| [24] | E. Shchepakina and V. Sobolev, Invariant surfaces of variable stability, J. Phys. Conf. Ser., 727 (2016), 012016. |
| [25] | V. Sobolev, Canard Cascades, Discrete Cont. Dyn-S, 18 (2013), 513-521. |
| [26] | A. Korobeinikov, E. Shchepakina and V. Sobolev, Paradox of enrichment and system order reduction: bacteriophages dynamics as case study, Math. Med. Biol., 33 (2016), 359-369. |
| [27] | M. P. Mortell, R. E. O'Malley, A. Pokrovskii, et al. (eds): Singular Perturbation and Hysteresis, SIAM, Philadelphia, 2005. |
| [28] | G. Ledder, Scaling for Dynamical Systems in Biology, Bull. Math. Biol., 79 (2017), 2747-2772. |
| [29] | M. Rosenzweig, The paradox of enrichment, Science, 171 (1971), 385-387. |
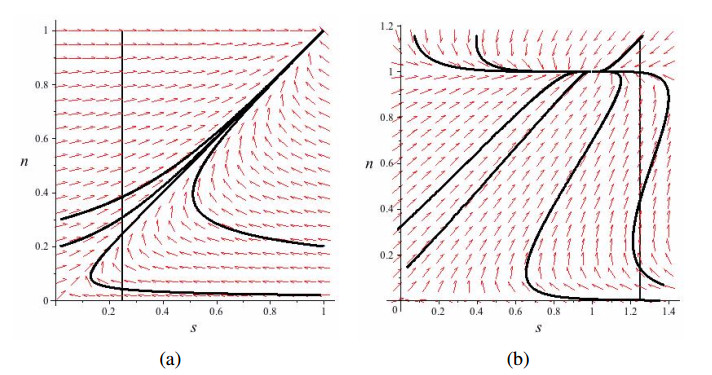
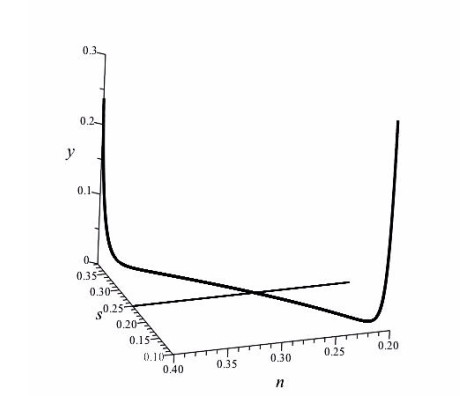
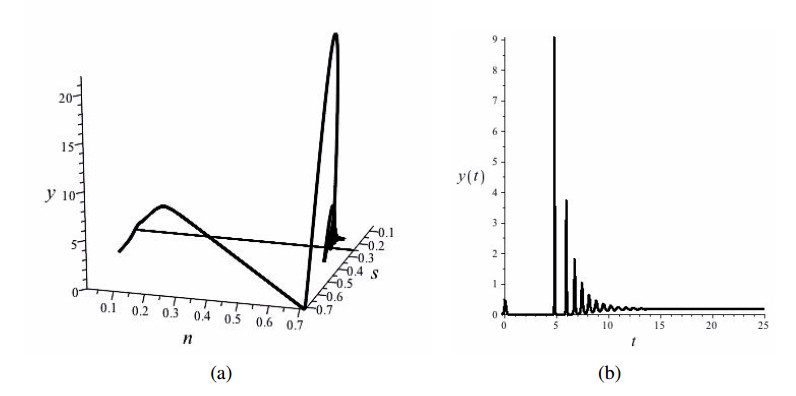
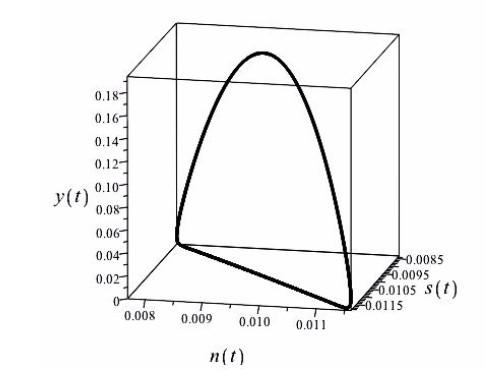
Figures(4)

Andrei Korobeinikov, Elena Shchepakina, Vladimir Sobolev. A black swan and canard cascades in an SIR infectious disease model[J]. Mathematical Biosciences and Engineering, 2020, 17(1): 725-736. doi: 10.3934/mbe.2020037







 DownLoad:
DownLoad: