Citation: Alina Chertock, Pierre Degond, Sophie Hecht, Jean-Paul Vincent. Incompressible limit of a continuum model of tissue growth with segregation for two cell populations[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 5804-5835. doi: 10.3934/mbe.2019290

| [1] | F. Otto, The geometry of dissipative evolution equations: the porous medium equation, Comm. Part. Diff. Eq., 26(2001), 101–174, URL https://doi.org/10.1081/PDE-100002243. |
| [2] | D. Kinderlehrer and N. J. Walkington, Approximation of parabolic equations using the Wasser-stein metric, ESAIM: M2AN, 33(1999), 837–852, URL https://doi.org/10.1051/m2an: 1999166. |
| [3] | R. Jordan, D. Kinderlehrer and F. Otto, The variational formulation of the Fokker–Planck equation, SIAM J. Math. Anal., 29(1998), 1–17, URL https://doi.org/10.1137/S0036141096303359. |
| [4] | D. Drasdo and S. Höhme, A single-cell-based model of tumor growth in vitro: Monolayers and spheroids, Phys. Biol., 2(2015), 133–147. |
| [5] | J. Galle, M. Loeffler and D. Drasdo, Modeling the effect of deregulated proliferation and apoptosis on the growth dynamics of epithelial cell populations in vitro, Biophys. J., 88(2005), 62–75, URL http://www.sciencedirect.com/science/article/pii/S0006349505730873. |
| [6] | R. Araujo and D. McElwain, A history of the study of solid tumour growth: the contribution of mathematical modelling, D.L.S. Bull. Math. Biol., 66(2004), 1039. |
| [7] | D. Bresch, T. Colin, E. Grenier, et al., Computational modeling of solid tumor growth: the avascular stage, SIAM J. Sci. Comput., 32(2010), 2321–2344, URL https://doi.org/10.1137/070708895. |
| [8] | H. Byrne and M. Chaplain, Growth of necrotic tumors in the presence and absence of inhibitors, Math. Biosci., 135(1996), 187–216, URL http://www.sciencedirect.com/science/article/pii/0025556496000235. |
| [9] | P. Ciarletta, L. Foret and M. Ben Amar, The radial growth phase of malignant melanoma: multi-phase modelling, numerical simulations and linear stability analysis, J. Royal Soc. Interface, 8(2011), 345–368, URL http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3030817/. |
| [10] | J. Ranft, M. Basan, J. Elgeti, et al., Fluidization of tissues by cell division and apoptosis, Proc. Natl. Acad. Sci., 107(2010), 20863–20868. |
| [11] | S. Cui and J. Escher, Asymptotic behaviour of solutions of a multidimensional moving boundary problem modeling tumor growth, Comm. Part. Diff. Eq., 33(2008), 636–655, URL https://doi.org/10.1080/03605300701743848. |
| [12] | A. Friedman and B. Hu, Stability and instability of Liapunov-Schmidt and Hopf bifurcation for a free boundary problem arising in a tumor model, Trans. Am. Math. Soc., 360(2008), 5291–5342. |
| [13] | H. P. Greenspan, Models for the growth of a solid tumor by diffusion, Stud. Appl. Math., 51(1972), 317–340, URL http://dx.doi.org/10.1002/sapm1972514317. |
| [14] | J. S. Lowengrub, H. B. Frieboes, F. Jin, et al., Nonlinear modelling of cancer: bridging the gap between cells and tumours, Nonlinearity, 23(2010), R1–R9, URL http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2929802/. |
| [15] | D. Hilhorst, M. Mimura and R. Schätzle, Vanishing latent heat limit in a Stefan-like problem aris-ing in biology, Nonlinear Anal. Real, 4(2003), 261 – 285, URL http://www.sciencedirect.com/science/article/pii/S1468121802000093. |
| [16] | B. Perthame, F. Quirós and J. L. Vázquez, The Hele–Shaw asymptotics for mechanical models of tumor growth, Arch. Ration. Mech. Anal., 212(2014), 93–127, URL https://doi.org/10.1007/s00205-013-0704-y. |
| [17] | B. Perthame, F. Quiròs, M. Tang et al., Derivation of a Hele-Shaw type system from a cell model with active motion, Interfaces Free Bound., 14(2014), 489–508. |
| [18] | B. Perthame and N. Vauchelet, Incompressible limit of a mechanical model of tumour growth with viscosity, Philos. Trans. R. Soc. A-Math. Phys. Eng. Sci., 373(2015), 20140283, URL http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4535270/. |
| [19] | H. Byrne and D. Drasdo, Individual-based and continuum models of growing cell popu-lations: a comparison, J. Math. Biol., 58(2008), 657, URL https://doi.org/10.1007/s00285-008-0212-0. |
| [20] | J. L. Vazquez, The Porous Medium Equation: Mathematical Theory, Oxford Mathematical Monographs, 2007. |
| [21] | S. Hecht and N. Vauchelet, Incompressible limit of a mechanical model for tissue growth with non-overlapping constraint, Commun. Math. Sci., 15(2017), 1913–1932, URL http://www.ncbi.nlm.nih.gov/pmc/articles/PMC5669502/. |
| [22] | A. Friedman and S. Y. Huang, Asymptotic behavior of solutions of ut = ∆φm(u) as m → ∞ with inconsistent initial values, Analyse Math. Appl., (1988), 165–180. |
| [23] | P. Bénilan, L. Boccardo and M. Herrero, On the limit of solutions of ut = ∆um as m → ∞., Interfaces Free Bound., 12(1989). |
| [24] | P. Bénilan and N. Igbida, La limite de la solution de ut = ∆pum lorsque m → ∞., C. R. Acad. Sci. Paris Sier, 321(1995), 1323–1328. |
| [25] | C. Elliott, M. Herrero, J. King, et al., The mesa problem: diffusion patterns for u t = ∇(u m ∇u) as m → ∞, IMA J. Appl. Math., 2(1986), 147–154. |
| [26] | O. Gil and F. Quirós, Convergence of the porous media equation to Hele-Shaw, Nonlinear Anal., 44(2001), 1111–1131, URL http://dx.doi.org/10.1016/S0362-546X(99)00325-9. |
| [27] | A. K. Dutt, Turing pattern amplitude equation for a model glycolytic reaction-diffusion system, J. Math. Chem., 48(2010), 841–855, URL https://doi.org/10.1007/s10910-010-9699-x. |
| [28] | E. F. Keller and L. A. Segel, Initiation of slime mold aggregation viewed as an instability, J. The-oret. Biol., 26(1970), 399–415, URL http://www.sciencedirect.com/science/article/pii/0022519370900925. |
| [29] | A. J. Lotka, Contribution to the theory of periodic reactions, J. Phys. Chem., 14(1909), 271–274, URL http://dx.doi.org/10.1021/j150111a004. |
| [30] | M. Bertsch, M. E. Gurtin and L. A. P. D. Hilhorst, On interacting populations that disperse to avoid crowding: the effect of a sedentary colony, Q. Appl. Math., 19(1984), 1–12. |
| [31] | S. N. Busenberg and C. C. Travis, Epidemic models with spatial spread due to population migra-tion, J. Math. Biol., 16(1983), 181–198, URL https://doi.org/10.1007/BF00276056. |
| [32] | M. Bertsch, R. D. Passo and M. Mimura, A free boundary problem arising in a simplified tumour growth model of contact inhibition, Interfaces Free Bound., 12(2010), 235–250. |
| [33] | M. Bertsch, D. Hilhorst, H. Izuhara, et al., A non linear parabolic-hyperbolic system for contact inhibition of cell growth, Differ. Equ. Appl., 4(2010), 137–157. |
| [34] | A. J. Sherratt and M. Chaplain, A new mathematical model for avascular tumour growth, J. Math. Biol., 43(2001), 291–312. |
| [35] | G. Galiano, S. Shmarev and J. Velasco, Existence and multiplicity of segregated solutions to a cell-growth contact inhibition problem, Discrete Contin. Dyn. Syst., 96(2015), 339–357, URL http://aimsciences.org/journals/displayArticlesnew.jsp?paperID=10564. |
| [36] | C. Elliott and Z. Songmu, On the Cahn-Hilliard equation, Ration. Mech. Anal., 35(1986), 1479–1501. |
| [37] | J. A. Carrillo, S. Fagioli, F. Santambrogio, et al., Splitting schemes and segregation in reaction cross-diffusion systems, J. Math. Anal., 50(2017),5695–5718. |
| [38] | I. Kim and A. R. Mészáros, On nonlinear cross-diffusion systems: an optimal transport approach, Calc. Var. Partial Diff. Eq., 57 (2018), 79, URL https://doi.org/10.1007/s00526-018-1351-9. |
| [39] | A. Chertock, P. Degond and J. Neusser, An asymptotic-preserving method for a relaxation of the Navier–Stokes–Korteweg equations, J. Comput. Phys., 335(2017), 387 – 403, URL http://www.sciencedirect.com/science/article/pii/S0021999117300463. |
| [40] | C. Rohde, On local and non-local Navier-Stokes-Korteweg systems for liquid-vapour phase tran-sitions, ZAMM Z. Angew. Math. Mech, 85(2005), 839–857. |
| [41] | J. A. Carrillo, A. Chertock and Y. Huang, A finite-volume method for nonlinear nonlocal equations with a gradient flow structure, Commun. Comput. Phys., 17(2015), 233–258. |
| [42] | S. Gottlieb, D. Ketcheson and C. W. Shu, Strong stability preserving Runge-Kutta and multistep time discretizations, World Scientific Publishing Co. Pte. Ltd. (2011). |
| [43] | S. Gottlieb, C. W. Shu and E. Tadmor, Strong stability-preserving high-order time discretization methods, SIAM Rev., 43(2001), 89–112. |
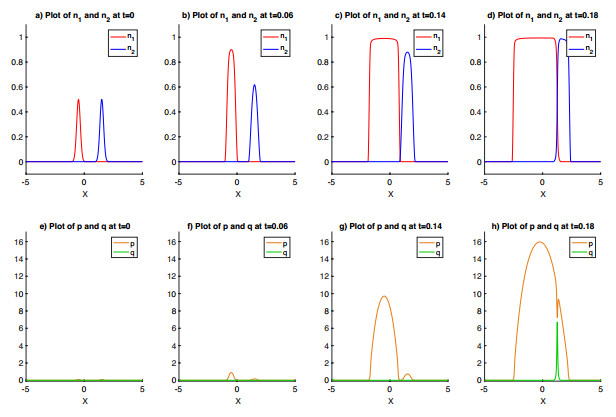
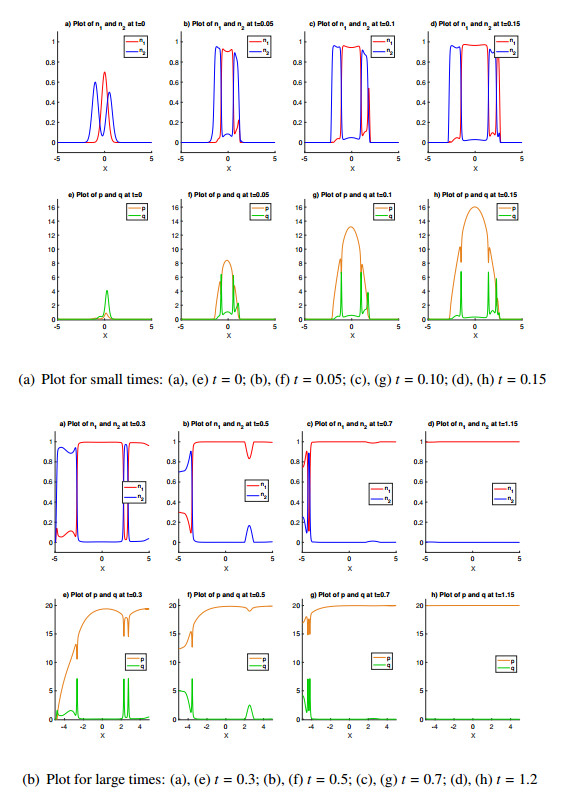
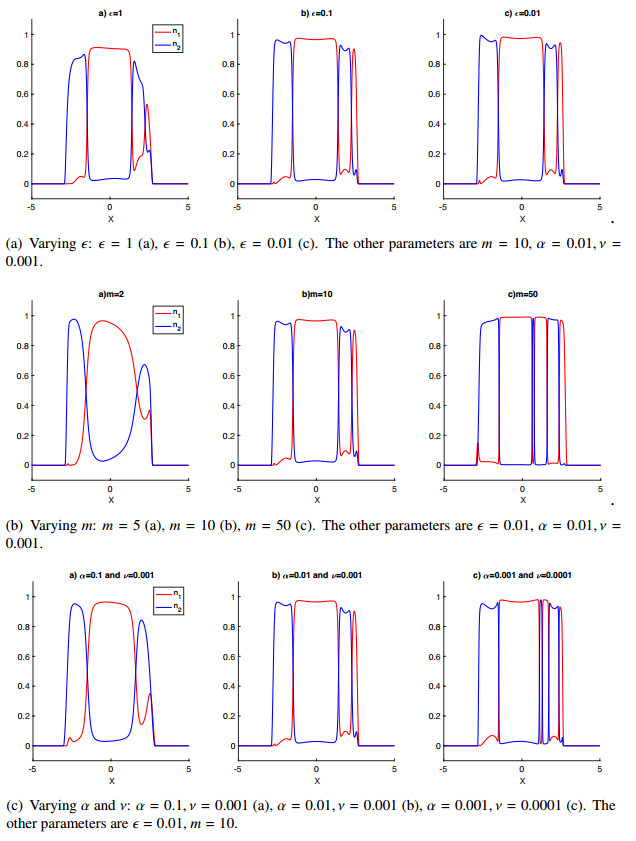
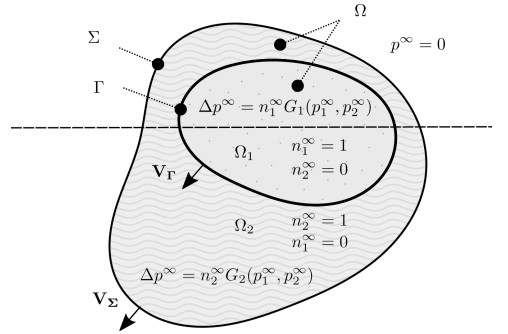

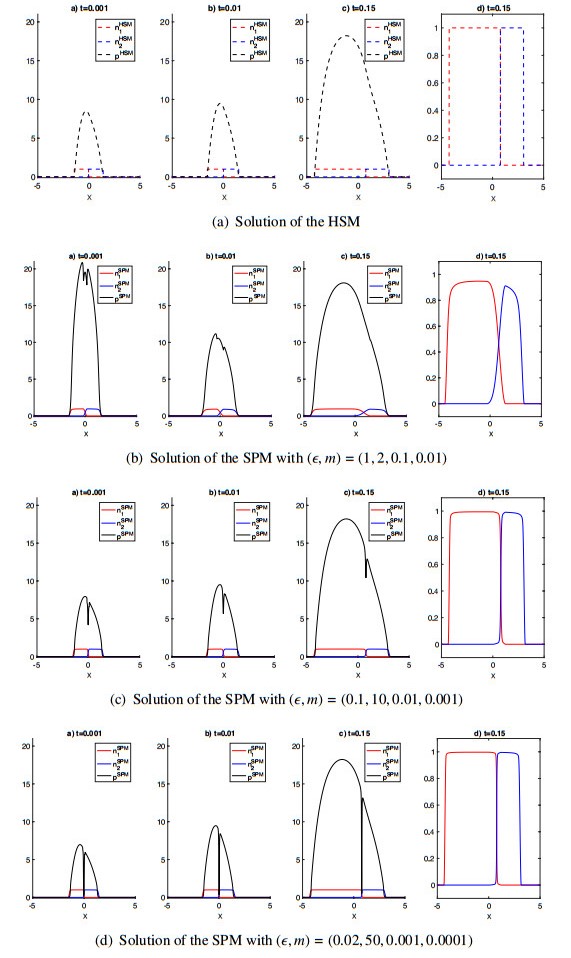
Figures(6)

Alina Chertock, Pierre Degond, Sophie Hecht, Jean-Paul Vincent. Incompressible limit of a continuum model of tissue growth with segregation for two cell populations[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 5804-5835. doi: 10.3934/mbe.2019290







 DownLoad:
DownLoad: