Citation: Zhan Chen, Yuting Zou. A multiscale model for heterogeneous tumor spheroid in vitro[J]. Mathematical Biosciences and Engineering, 2018, 15(2): 361-392. doi: 10.3934/mbe.2018016

| [1] | [ S. Aland,H. Hatzikirou,J. Lowengrub,A. Voigt, A mechanistic collective cell model for epithelial colony growth and contact inhibition, Biophysical Journal, 109 (2015): 1347-1357. |
| [2] | [ R. K. Banerjee,W. W. van Osdol,P. M. Bungay,C. Sung,R. L. Dedrick, Finite element model of antibody penetration in a prevascular tumor nodule embedded in normal tissue, Journal of Controlled Release, 74 (2001): 193-202. |
| [3] | [ S. Breslin,L. O'Driscoll, Three-dimensional cell culture: The missing link in drug discovery, Drug Discovery Today, 18 (2013): 240-249. |
| [4] | [ G. W. Brodland, Computational modeling of cell sorting, tissue engulfment, and related phenomena: A review, Applied Mechanics Reviews, 57 (2004): 47-76. |
| [5] | [ G. W. Brodland,D. Viens,J. H. Veldhuis, A new cell-based fe model for the mechanics of embryonic epithelia, Computer Methods in Biomechanics and Biomedical Engineering, 10 (2007): 121-128. |
| [6] | [ J. C. Butcher, Numerical Methods for Ordinary Differential Equations John Wiley & Sons, 2016. |
| [7] | [ L. L. Campbell,K. Polyak, Breast tumor heterogeneity: Cancer stem cells or clonal evolution?, Cell Cycle, 6 (2007): 2332-2338. |
| [8] | [ J. Casciari,S. Sotirchos,R. Sutherland, Mathematical modelling of microenvironment and growth in emt6/ro multicellular tumour spheroids, Cell Proliferation, 25 (1992): 1-22. |
| [9] | [ J. Casciari,S. Sotirchos,R. Sutherland, Variations in tumor cell growth rates and metabolism with oxygen concentration, glucose concentration, and extracellular ph, Journal of Cellular Physiology, 151 (1992): 386-394. |
| [10] | [ P. Cirri,P. Chiarugi, Cancer-associated-fibroblasts and tumour cells: A diabolic liaison driving cancer progression, Cancer and Metastasis Reviews, 31 (2012): 195-208. |
| [11] | [ J. C. Dallon,H. G. Othmer, How cellular movement determines the collective force generated by the Dictyostelium discoideum slug, J. Theor. Biol., 231 (2004): 203-222. |
| [12] | [ T. S. Deisboeck,Z. Wang,P. Macklin,V. Cristini, Multiscale cancer modeling, Ann. Rev. Biomed. Eng., 13 (2011): 127-155. |
| [13] | [ M. J. Dorie,R. F. Kallman,M. A. Coyne, Effect of cytochalasin b, nocodazole and irradiation on migration and internalization of cells and microspheres in tumor cell spheroids, Experimental Cell Research, 166 (1986): 370-378. |
| [14] | [ M. J. Dorie,R. F. Kallman,D. F. Rapacchietta,D. Van Antwerp,Y. R. Huang, Migration and internalization of cells and polystyrene microspheres in tumor cell spheroids, Experimental Cell Research, 141 (1982): 201-209. |
| [15] | [ D. Drasdo,S. Höhme, A single-cell-based model of tumor growth in vitro: Monolayers and spheroids, Physical Biology, 2 (2005): 133-147. |
| [16] | [ D. Duguay,R. A. Foty,M. S. Steinberg, Cadherin-mediated cell adhesion and tissue segregation: Qualitative and quantitative determinants, Developmental Biology, 253 (2003): 309-323. |
| [17] | [ K. Erbertseder, J. Reichold, B. Flemisch, P. Jenny and R. Helmig, A coupled discrete/continuum model for describing cancer-therapeutic transport in the lung PloS One 7 (2012), e31966. |
| [18] | [ E. Evans, Detailed mechanics of membrane-membrane adhesion and separation. ii. discrete kinetically trapped molecular cross-bridges, Biophysical Journal, 48 (1985): 185-192. |
| [19] | [ E. A. Evans, Detailed mechanics of membrane-membrane adhesion and separation. i. continuum of molecular cross-bridges, Biophysical Journal, 48 (1985): 175-183. |
| [20] | [ E. M. Felipe De Sousa,L. Vermeulen,E. Fessler,J. P. Medema, Cancer heterogeneity-a multifaceted view, EMBO Reports, 14 (2013): 686-695. |
| [21] | [ T. Fiaschi and P. Chiarugi, Oxidative stress, tumor microenvironment, and metabolic reprogramming: A diabolic liaison International Journal of Cell Biology 2012 (2012), Article ID 762825, 8pp. |
| [22] | [ R. A. Foty,M. S. Steinberg, Cadherin-mediated cell-cell adhesion and tissue segregation in relation to malignancy, International Journal of Developmental Biology, 48 (2004): 397-409. |
| [23] | [ R. A. Foty,M. S. Steinberg, The differential adhesion hypothesis: A direct evaluation, Developmental Biology, 278 (2005): 255-263. |
| [24] | [ R. A. Foty,M. S. Steinberg, Differential adhesion in model systems, Wiley Interdisciplinary Reviews: Developmental Biology, 2 (2013): 631-645. |
| [25] | [ J. Freyer,R. Sutherland, A reduction in the in situ rates of oxygen and glucose consumption of cells in emt6/ro spheroids during growth, Journal of Cellular Physiology, 124 (1985): 516-524. |
| [26] | [ J. Galle,G. Aust,G. Schaller,T. Beyer,D. Drasdo, Individual cell-based models of the spatial-temporal organization of multicellular systems-achievements and limitations, Cytometry Part A, 69 (2006): 704-710. |
| [27] | [ D. Garrod,M. Steinberg, Tissue-specific sorting-out in two dimensions in relation to contact inhibition of cell movement, Nature, 244 (1973): 568-569. |
| [28] | [ P. Gerlee,A. R. Anderson, An evolutionary hybrid cellular automaton model of solid tumour growth, Journal of Theoretical Biology, 246 (2007): 583-603. |
| [29] | [ M. Gerlinger,A. J. Rowan,S. Horswell,J. Larkin,D. Endesfelder,E. Gronroos,P. Martinez,N. Matthews,A. Stewart,P. Tarpey, Intratumor heterogeneity and branched evolution revealed by multiregion sequencing, New England Journal of Medicine, 366 (2012): 883-892. |
| [30] | [ R. H. Grantab and I. F. Tannock, Penetration of anticancer drugs through tumour tissue as a function of cellular packing density and interstitial fluid pressure and its modification by bortezomib BMC Cancer 12 (2012), 214. |
| [31] | [ J. B. Green, Sophistications of cell sorting, Nature Cell Biology, 10 (2008): 375-377. |
| [32] | [ E. Hairer, S. Norsett and G. Wanner, Solving Ordinary Differential Equations I: Nonstiff Problems, Second edition. Springer Series in Computational Mathematics, 8. Springer-Verlag, Berlin, 1993. |
| [33] | [ J. W. Haycock, 3d cell culture: A review of current approaches and techniques, 3D Cell Culture, 695 (2010): 1-15. |
| [34] | [ G. Helmlinger,P. A. Netti,H. C. Lichtenbeld,R. J. Melder,R. K. Jain, Solid stress inhibits the growth of multicellular tumor spheroids, Nature Biotechnology, 15 (1997): 778-783. |
| [35] | [ F. Hirschhaeuser,H. Menne,C. Dittfeld,J. West,W. Mueller-Klieser,L. A. Kunz-Schughart, Multicellular tumor spheroids: An underestimated tool is catching up again, Journal of Biotechnology, 148 (2010): 3-15. |
| [36] | [ M. S. Hutson, G. W. Brodland, J. Yang and D. Viens, Cell sorting in three dimensions: Topology, fluctuations, and fluidlike instabilities Physical Review Letters 101 (2008), 148105. |
| [37] | [ J. N. Jennings, A New Computational Model for Multi-cellular Biological Systems PhD thesis, University of Cambridge, 2014. |
| [38] | [ Y. Jiang,H. Levine,J. Glazier, Possible cooperation of differential adhesion and chemotaxis in mound formation of dictyostelium, Biophysical Journal, 75 (1998): 2615-2625. |
| [39] | [ Y. Jiang,J. Pjesivac-Grbovic,C. Cantrell,J. P. Freyer, A multiscale model for avascular tumor growth, Biophysical journal, 89 (2005): 3884-3894. |
| [40] | [ K. Kendall, Adhesion: Molecules and mechanics, Science, 263 (1994): 1720-1725. |
| [41] | [ Z. I. Khamis, Z. J. Sahab and Q. X. A. Sang, Active roles of tumor stroma in breast cancer metastasis International Journal of Breast Cancer 2012 (2012), Article ID 574025, 10pp. |
| [42] | [ Y. Kim,M. Stolarska,H. Othmer, The role of the microenvironment in tumor growth and invasion, Progress in Biophysics and Molecular Biology, 106 (2011): 353-379. |
| [43] | [ Y. Kim,H. G. Othmer, A hybrid model of tumor-stromal interactions in breast cancer, Bull. Math. Biol., 75 (2013): 1304-1350. |
| [44] | [ Y. KIM,S. ROH, A hybrid model for cell proliferation and migration in glioblastoma, Discrete & Continuous Dynamical Systems-Series B, 18 (2013): 969-1015. |
| [45] | [ Y. Kim,M. A. Stolarska,H. G. Othmer, A hybrid model for tumor spheroid growth in vitro i: Theoretical development and early results, Mathematical Models and Methods in Applied Sciences, 17 (2007): 1773-1798. |
| [46] | [ L. C. Kimlin,G. Casagrande,V. M. Virador, In vitro three-dimensional (3d) models in cancer research: An update, Molecular Carcinogenesis, 52 (2013): 167-182. |
| [47] | [ T. Lecuit,P.-F. Lenne, Cell surface mechanics and the control of cell shape, tissue patterns and morphogenesis, Nature Reviews Molecular Cell Biology, 8 (2007): 633-644. |
| [48] | [ X.-F. Li,S. Carlin,M. Urano,J. Russell,C. C. Ling,J. A. O'Donoghue, Visualization of hypoxia in microscopic tumors by immunofluorescent microscopy, Cancer Research, 67 (2007): 7646-7653. |
| [49] | [ D. Loessner,J. P. Little,G. J. Pettet,D. W. Hutmacher, A multiscale road map of cancer spheroids-incorporating experimental and mathematical modelling to understand cancer progression, J Cell Sci, 126 (2013): 2761-2771. |
| [50] | [ P. Macklin,S. McDougall,A. R. Anderson,M. A. Chaplain,V. Cristini,J. Lowengrub, Multiscale modelling and nonlinear simulation of vascular tumour growth, Journal of Mathematical Biology, 58 (2009): 765-798. |
| [51] | [ J.-L. Maître,H. Berthoumieux,S. F. G. Krens,G. Salbreux,F. Jülicher,E. Paluch,C.-P. Heisenberg, Adhesion functions in cell sorting by mechanically coupling the cortices of adhering cells, Science, 338 (2012): 253-256. |
| [52] | [ M. Martins,S. Ferreira,M. Vilela, Multiscale models for the growth of avascular tumors, Physics of Life Reviews, 4 (2007): 128-156. |
| [53] | [ A. Marusyk,V. Almendro,K. Polyak, Intra-tumour heterogeneity: A looking glass for cancer?, Nature Reviews Cancer, 12 (2012): 323-334. |
| [54] | [ D. McElwain,G. Pettet, Cell migration in multicell spheroids: Swimming against the tide, Bulletin of Mathematical Biology, 55 (1993): 655-674. |
| [55] | [ E. Méhes, E. Mones, V. Németh and T. Vicsek, Collective motion of cells mediates segregation and pattern formation in co-cultures, PloS One 7. |
| [56] | [ L. M. F. Merlo,J. W. Pepper,B. J. Reid,C. C. Maley, Cancer as an evolutionary and ecological process, Nature Reviews Cancer, 6 (2006): 924-935. |
| [57] | [ D. Miller, Sugar uptake as a function of cell volume in human erythrocytes, The Journal of Physiology, 170 (1964): 219-225. |
| [58] | [ W. F. Mueller-Klieser,R. M. Sutherland, Oxygen consumption and oxygen diffusion properties of multicellular spheroids from two different cell lines, in Oxygen Transport to Tissue-VI , Springer, 180 (1984): 311-321. |
| [59] | [ S. M. Mumenthaler,J. Foo,N. C. Choi,N. Heise,K. Leder,D. B. Agus,W. Pao,F. Michor,P. Mallick, The impact of microenvironmental heterogeneity on the evolution of drug resistance in cancer cells, Cancer Informatics, 14 (2015): 19-31. |
| [60] | [ S. Mumenthaler,J. Foo,K. Leder,N. Choi,D. Agus,W. Pao,P. Mallick,F. Michor, Evolutionary modeling of combination treatment strategies to overcome resistance to tyrosine kinase inhibitors in non-small cell lung cancer, Molecular Pharmaceutics, 8 (2011): 2069-2079. |
| [61] | [ T. J. Newman, Modeling multi-cellular systems using sub-cellular elements, Math. Biosci. Eng., 2 (2005), 613–624, arXiv preprint q-bio/0504028. |
| [62] | [ H. Ninomiya,R. David,E. W. Damm,F. Fagotto,C. M. Niessen,R. Winklbauer, Cadherin-dependent differential cell adhesion in xenopus causes cell sorting in vitro but not in the embryo, Journal of Cell Science, 125 (2012): 1877-1883. |
| [63] | [ E. Palsson, A three-dimensional model of cell movement in multicellular systems, Future Generation Computer Systems, 17 (2001): 835-852. |
| [64] | [ E. Palsson, A 3-d model used to explore how cell adhesion and stiffness affect cell sorting and movement in multicellular systems, Journal of Theoretical Biology, 254 (2008): 1-13. |
| [65] | [ E. Palsson,H. G. Othmer, A model for individual and collective cell movement in dictyostelium discoideum, Proceedings of the National Academy of Sciences, 97 (2000): 10448-10453. |
| [66] | [ G. Pettet,C. Please,M. Tindall,D. McElwain, The migration of cells in multicell tumor spheroids, Bulletin of Mathematical Biology, 63 (2001): 231-257. |
| [67] | [ K. Polyak, Heterogeneity in breast cancer, The Journal of Clinical Investigation 121 (2011), 3786. |
| [68] | [ N. J. Poplawski,U. Agero,J. S. Gens,M. Swat,J. A. Glazier,A. R. Anderson, Front instabilities and invasiveness of simulated avascular tumors, Bulletin of Mathematical Biology, 71 (2009): 1189-1227. |
| [69] | [ A. Quarteroni, R. Sacco and F. Saleri, Matematica Numerica Springer Science & Business Media, 1998. |
| [70] | [ A. A. Qutub,F. M. Gabhann,E. D. Karagiannis,P. Vempati,A. S. Popel, Multiscale models of angiogenesis, Engineering in Medicine and Biology Magazine, IEEE, 28 (2009): 14-31. |
| [71] | [ K. A. Rejniak,R. H. Dillon, A single cell-based model of the ductal tumour microarchitecture, Computational and Mathematical Methods in Medicine, 8 (2007): 51-69. |
| [72] | [ T. Roose,P. A. Netti,L. L. Munn,Y. Boucher,R. K. Jain, Solid stress generated by spheroid growth estimated using a linear poroelastisity model, Microvascular Research, 66 (2003): 204-212. |
| [73] | [ G. Schaller and M. Meyer-Hermann, Multicellular tumor spheroid in an off-lattice voronoi-delaunay cell model Physical Review E 71 (2005), 051910, 16pp. |
| [74] | [ G. Schaller,M. Meyer-Hermann, Continuum versus discrete model: a comparison for multicellular tumour spheroids, Philosophical Transactions of the Royal Society of London A: Mathematical, Physical and Engineering Sciences, 364 (2006): 1443-1464. |
| [75] | [ E.-M. Schötz,R. D. Burdine,F. Jülicher,M. S. Steinberg,C.-P. Heisenberg,R. A. Foty, Quantitative differences in tissue surface tension influence zebrafish germ layer positioning, HFSP journal, 2 (2008): 42-56. |
| [76] | [ R. Shipley,S. Chapman, Multiscale modelling of fluid and drug transport in vascular tumours, Bulletin of Mathematical Biology, 72 (2010): 1464-1491. |
| [77] | [ A. Shirinifard, J. S. Gens, B. L. Zaitlen, N. J. Poplawski, M. Swat and J. A. Glazier, 3d multi-cell simulation of tumor growth and angiogenesis PloS One 4 (2009), e7190. |
| [78] | [ K. Smalley,M. Lioni,M. Herlyn, Life ins't flat: Taking cancer biology to the next dimension, In Vitro Cellular & Developmental Biology-Animal, 42 (2006): 242-247. |
| [79] | [ A. Starzec,D. Briane,M. Kraemer,J.-C. Kouyoumdjian,J.-L. Moretti,R. Beaupain,O. Oudar, Spatial organization of three-dimensional cocultures of adriamycin-sensitive and-resistant human breast cancer mcf-7 cells, Biology of the Cell, 95 (2003): 257-264. |
| [80] | [ M. S. Steinberg, Reconstruction of tissues by dissociated cells, Science, 141 (1963): 401-408. |
| [81] | [ M. S. Steinberg, Adhesion in development: An historical overview, Developmental Biology, 180 (1996): 377-388. |
| [82] | [ M. Steinberg,D. Garrod, Observations on the sorting-out of embryonic cells in monolayer culture, Journal of Cell Science, 18 (1975): 385-403. |
| [83] | [ M. A. Stolarska,Y. Kim,H. G. Othmer, Multi-scale models of cell and tissue dynamics, Philosophical Transactions of the Royal Society of London A: Mathematical, Physical and Engineering Sciences, 367 (2009): 3525-3553. |
| [84] | [ K. Sung,C. Dong,G. Schmid-Schönbein,S. Chien,R. Skalak, Leukocyte relaxation properties, Biophysical Journal, 54 (1988): 331-336. |
| [85] | [ M. H. Swat, S. D. Hester, R. W. Heiland, B. L. Zaitlen, J. A. Glazier and A. Shirinifard, Compucell3d manual and tutorial version 3. 5. 0. |
| [86] | [ G. Taraboletti,D. D. Roberts,L. A. Liotta, Thrombospondin-induced tumor cell migration: Haptotaxis and chemotaxis are mediated by different molecular domains, The Journal of Cell Biology, 105 (1987): 2409-2415. |
| [87] | [ K. Thompson,H. Byrne, Modelling the internalization of labelled cells in tumour spheroids, Bulletin of Mathematical Biology, 61 (1999): 601-623. |
| [88] | [ P. L. Townes,J. Holtfreter, Directed movements and selective adhesion of embryonic amphibian cells, Journal of Experimental Zoology, 128 (1955): 53-120. |
| [89] | [ G. Wayne Brodland,H. H. Chen, The mechanics of cell sorting and envelopment, Journal of Biomechanics, 33 (2000): 845-851. |
| [90] | [ D. G. Wilkinson, How attraction turns to repulsion, Nature Cell Biology, 5 (2003): 851-853. |
| [91] | [ M. Zanoni, F. Piccinini, C. Arienti, A. Zamagni, S. Santi, R. Polico, A. Bevilacqua and A. Tesei, 3d tumor spheroid models for in vitro therapeutic screening: A systematic approach to enhance the biological relevance of data obtained Scientific Reports 6 (2016), 19103. |
| [92] | [ Y. Zhang, G. Thomas, M. Swat, A. Shirinifard and J. Glazier, Computer simulations of cell sorting due to differential adhesion PloS One 6 (2011), e24999. |
| [93] | [ M. Zimmermann,C. Box,S. A. Eccles, Two-dimensional vs. three-dimensional in vitro tumor migration and invasion assays, in Target Identification and Validation in Drug Discovery, Springer, null (2013): 227-252. |
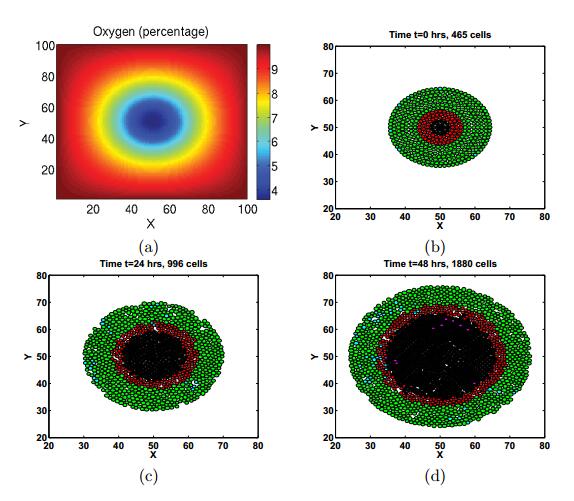
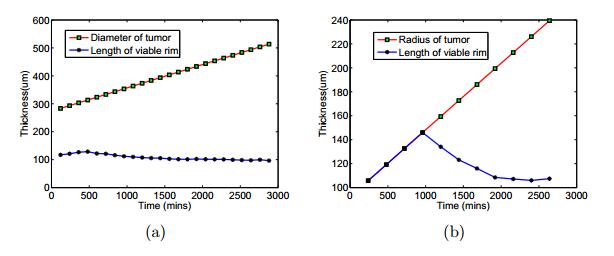

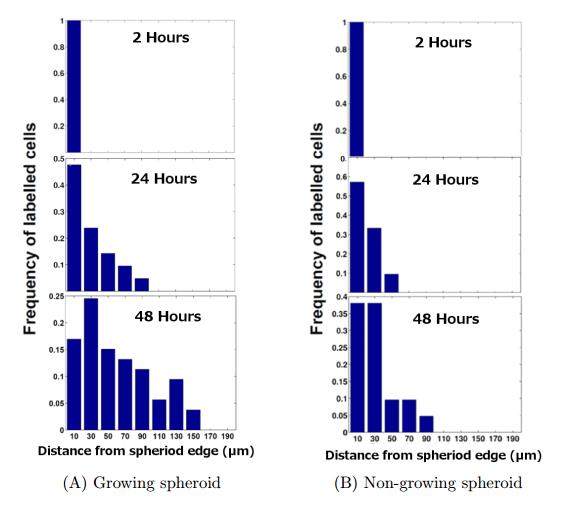
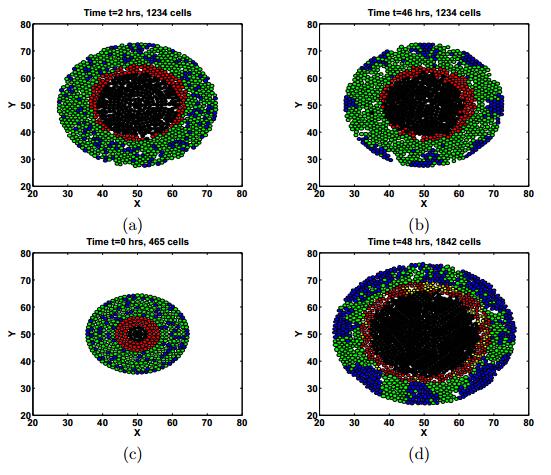
Figures(11) / Tables(3)

Zhan Chen, Yuting Zou. A multiscale model for heterogeneous tumor spheroid in vitro[J]. Mathematical Biosciences and Engineering, 2018, 15(2): 361-392. doi: 10.3934/mbe.2018016







 DownLoad:
DownLoad: