Citation: Hongyong Zhao, Qianjin Zhang, Linhe Zhu. The spatial dynamics of a zebrafish model with cross-diffusions[J]. Mathematical Biosciences and Engineering, 2017, 14(4): 1035-1054. doi: 10.3934/mbe.2017054

| [1] | [ R. Asai,R. Taguchi,Y. Kume,M. Saito,S. Kondo, Zebrafish Leopard gene as a component of the putative reaction-diffusion system, Mech. Dev., 89 (1999): 87-92. |
| [2] | [ V. Dufiet,J. Boissonade, Dynamics of turing pattern monolayers close to onset, Phys. Rev. E., 53 (1996): 1-10. |
| [3] | [ M. Fras,M. Gosak, Spatiotemporal patterns provoked by environmental variability in a predator-prey model, Biosystems, 114 (2013): 172-177. |
| [4] | [ A. Gierer,H. Meinhardt, A theory of biological pattern formation, Kybernetika, 12 (1972): 30-39. |
| [5] | [ P. Gray,S. K. Scott, Autocatalytic reactions in the isothermal continuous stirred tank reactor, Chem.Eng. Sci., 39 (1984): 1087-1097. |
| [6] | [ G. H. Gunaratne,Q. Ouyang,H. L. Swinney, Pattern formation in the presence of symmetries, Phys. Rev. E., 50 (1994): 4-15. |
| [7] | [ O. Jensen,V. C. Pannbacker,G. Dewel,P. Borckmans, Subcritical transitions to Turing structures, Phys. Lett. A., 179 (1993): 91-96. |
| [8] | [ C. T. Klein,F. F. Seelig, Turing structures in a system with regulated gap-junctions, Biosystems, 35 (1995): 15-23. |
| [9] | [ S. Kondo, The reaction-diffusion system: A mechanism for autonomous pattern for-mation in the animal skin, Genes. Cells., 7 (2002): 535-541. |
| [10] | [ S. Kondo,R. Asia, A reaction-diffusion wave on the skin of the marine angelfish Pomacanthus, Nature, 376 (1995): 765-767. |
| [11] | [ S. Kondo,H. Shirota, Theoretical analysis of mechanisms that generate the pigmen-tation pattern of animals, Semin.Cell. Dev. Biol., 20 (2009): 82-89. |
| [12] | [ H. Meinhardt, Reaction-diffusion system in development, Appl. Math. Comput Appl. Math. Comput., 32 (1989): 103-135. |
| [13] | [ S. Miyazawa,M. Okamoto,S. Kondo, Blending of animal colour patterns by hybridiza-tion, Nat. Commun, 10 (2010): 1-6. |
| [14] | [ M. Nguyen,A.M. Stewart,A.V. Kalueff, Aquatic blues: Modeling depression and antide-pressant action in zebrafish, Prog. Neuro-Psychoph, 55 (2014): 26-39. |
| [15] | [ Ouyang Q., 2000. Pattern Formation in Reaction-diffusion Systems, (Shanghai: Shanghai Sci-Tech Education Publishing House)(in Chinese), |
| [16] | [ D.M. Parichy, Pigment patterns: Fish in stripes and spots current, Biology, 13 (2003): 947-950. |
| [17] | [ J. Schnackenberg, Simple chemical reaction systems with limit cycle behavior, J. Theor. Biol., 81 (1979): 389-400. |
| [18] | [ H. Shoji,Y. Iwasa, Labyrinthine versus straight-striped patterns generated by two-dimensional Turing systems, J.Theor. Biol., 237 (2005): 104-116. |
| [19] | [ H. Shoji,Y. Iwasa, Pattern selection and the direction of stripes in two-dimensional turing systems for skin pattern formation of fishes Formation of Fishes, Forma, 18 (2003): 3-18. |
| [20] | [ H. Shoji,Y. Iwasa,S. Kondo, Stripes, spots, or reversedspots in two-dimensional Turing systems, J. Theor. Biol., 224 (2003): 339-350. |
| [21] | [ H. Shoji,Y. Iwasa,A. Mochizuki,S. Kondo, Directionality of stripes formed by anisotropic reaction-diffusion models, J. Theor. Biol., 214 (2002): 549-561. |
| [22] | [ H. Shoji,A. Mochizuki,Y. Iwasa,M. Hirata,T. Watanabe,S. Hioki,S. Kondo, Origin of directionality in the fish stripe pattern, Dev. Dynam., 226 (2003): 627-633. |
| [23] | [ A.M. Stewart,E. Yang,M. Nguyen,A.V. Kalueff, Developing zebrafish models relevant to PTSD and other trauma-and stressor-related disorders, Prog. Neuro-Psychoph, 55 (2014): 67-79. |
| [24] | [ G. Q. Sun,Z. Jin,Q. X. Liu,B. L. Li, Rich dynamics in a predator-prey model with both noise and periodic force, Biosystems, 100 (2010): 14-22. |
| [25] | [ Wang, W. M. Liu, H. Y. Cai Y. L. and Liu, Z. Q. (2011), Turing pattern selection in a reaction diffusion epidemic model, Chin. Phys. B., 074702, 12pp. |
| [26] | [ M. Yamaguchi,E. Yoshimoto,S. Kondo, Pattern regulation in the stripe of zebrafish suggests an underlying dynamic and autonomous mechanism, PNAS, 104 (2007): 4790-4793. |
| [27] | [ X. Y. Yang,T. Q. Liu,J. J. Zhang,T. S. Zhou, The mechanism of Turing pattern formation in a positive feedback system with cross diffusion, J. Stat. Mech-Theory. E., 14 (2014): 1-16. |
| [28] | [ Zhang, X. C. Sun G. Q. and Jin, Z. Spatial dynamics in a predator-prey model with Beddington-DeAngelis functional response, Phys. Rev. E., (2012), 021924, 14pp. |
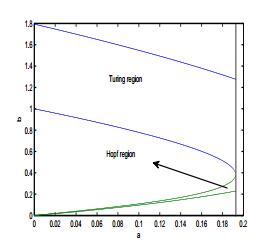
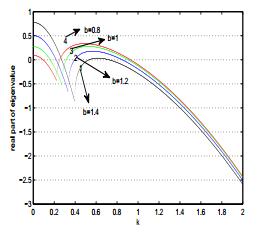
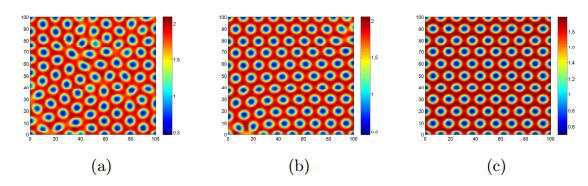

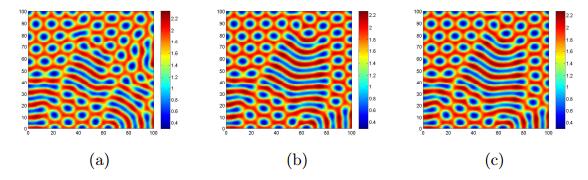




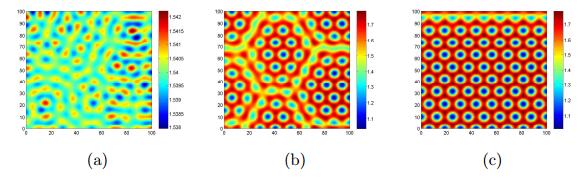


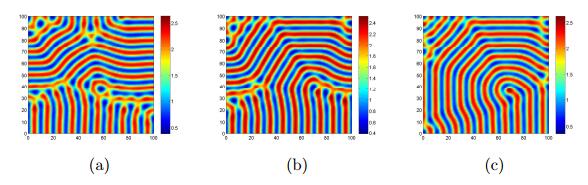
Figures(17)

Hongyong Zhao, Qianjin Zhang, Linhe Zhu. The spatial dynamics of a zebrafish model with cross-diffusions[J]. Mathematical Biosciences and Engineering, 2017, 14(4): 1035-1054. doi: 10.3934/mbe.2017054







 DownLoad:
DownLoad: