Citation: Youcef Mammeri, Damien Sellier. A surface model of nonlinear, non-steady-state phloem transport[J]. Mathematical Biosciences and Engineering, 2017, 14(4): 1055-1069. doi: 10.3934/mbe.2017055

| [1] | [ P. Cabrita,M. Thorpe,G. Huber, Hydrodynamics of steady state phloem transport with radial leakage of solute, Frontiers Plant Sci., 4 (2013): 531-543. |
| [2] | [ A. L. Christy,J. M. Ferrier, A mathematical treatment of Münch's pressure-flow hypothesis of phloem translocation, Plant Physio., 52 (1973): 531-538. |
| [3] | [ T. K. Dey,J. A. Levine, Delaunay meshing of isosurfaces, Visual Comput., 24 (2008): 411-422. |
| [4] | [ J. M. Ferrier, Further theoretical analysis of concentration-pressure-flux waves in phloem transport systems, Can. J. Bot., 56 (1978): 1086-1090. |
| [5] | [ F. G. Feugier,A. Satake, Dynamical feedback between circadian clock and sucrose availability explains adaptive response of starch metabolism to various photoperiods, Frontiers Plant Sci., 305 (2013): 1-11. |
| [6] | [ D. B. Fisher,C. Cash-Clark, Sieve tube unloading and post-phloemtransport of fluorescent tracers and proteins injected into sieve tubes via severed aphid stylets, Plant Physio., 123 (2000): 125-137. |
| [7] | [ J. D. Goeschl,C. E. Magnuson, Physiological implications of the Münch--Horwitz theory of phloem transport: effect of loading rates, Plant Cell Env., 9 (1986): 95-102. |
| [8] | [ J. Gričar,L. Krže,K. Čufar, Number of cells in xylem, phloem and dormant cambium in silver fir (Abies alba), in trees of different vitality, IAWA Journal, 30 (2009): 121-133. |
| [9] | [ J. Hansen,E. Beck, The fate and path of assimilation products in the stem of 8-year-old {Scots} pine (Pinus sylvestris {L}.) trees, Trees, 4 (1990): 16-21. |
| [10] | [ F. Hecht, New Developments in Freefem++, J. Num. Math., 20 (2012): 251-265. |
| [11] | [ L. Horwitz, Some simplified mathematical treatments of translocation in plants, Plant Physio., 33 (1958): 81-93. |
| [12] | [ T. Hölttä,M. Mencuccini,E. Nikinmaa, Linking phloem function to structure: Analysis with a coupled xylem-phloem transport model, J. Theo. Bio., 259 (2009): 325-337. |
| [13] | [ T. Hölttä,T. Vesala,S. Sevanto,M. Perämäki,E. Nikinmaa, Modeling xylem and phloem water flows in trees according to cohesion theory and Münch hypothesis, Trees, 20 (2006): 67-78. |
| [14] | [ W. Hundsdorfer and J. G. Verwer, Numerical Solution of Time-Dependent Advection-Diffusion-Reaction Equations Springer Series in Comput. Math., 33, Springer, 2003. |
| [15] | [ K. H. Jensen,J. Lee,T. Bohr,H. Bruus,N. M. Holbrook,M. A. Zwieniecki, Optimality of the Münch mechanism for translocation of sugars in plants, J. R. Soc. Interface, 8 (2011): 1155-1165. |
| [16] | [ A. Kagawa,A. Sugimoto,T. C. Maximov, CO 2 pulse-labelling of photoassimilates reveals carbon allocation within and between tree rings, Plant Cell Env., 29 (2006): 1571-1584. |
| [17] | [ E. M. Kramer, Wood grain pattern formation: A brief review, J. Plant Growth Reg., 25 (2006): 290-301. |
| [18] | [ H.-O. Kreiss and J. Lorenz, Initial-Boundary Value Problems and the Navier-Stokes Equations Classics in Applied Mathematics, SIAM, 2004. |
| [19] | [ A. Lacointe,P. E. H. Minchin, Modelling phloem and xylem transport within a complex architecture, Funct. Plant Bio., 35 (2008): 772-780. |
| [20] | [ A. Lang, A model of mass flow in the phloem, Funct. Plant Bio., 5 (1978): 535-546. |
| [21] | [ P. E. H. Minchin,M. R. Thorpe,J. F. Farrar, A simple mechanistic model of phloem transport which explains sink priority, Journal of Experimental Botany, 44 (1993): 947-955. |
| [22] | [ E. Münch, Die Stoffbewegungen in der Pflanze Jena, Gustav Fischer, 1930. |
| [23] | [ K. A. Nagel,B. Kastenholz,S. Jahnke,D. van Dusschoten,T. Aach,M. Mühlich,D. Truhn,H. Scharr,S. Terjung,A. Walter,U. Schurr, Temperature responses of roots: Impact on growth, root system architecture and implications for phenotyping, Funct. Plant Bio., 36 (2009): 947-959. |
| [24] | [ E. M. Ouhabaz, Analysis of Heat Equations on Domains London Math. Soc. Monographs Series, Princeton University Press, 2005. |
| [25] | [ S. Payvandi,K. R. Daly,K. C. Zygalakis,T. Roose, Mathematical modelling of the phloem: The importance of diffusion on sugar transport at osmotic equilibrium, Bull. Math Biol., 76 (2014): 2834-2865. |
| [26] | [ S. Pfautsch,J. Renard,M. G. Tjoelker,A. Salih, Phloem as capacitor: Radial transfer of water into xylem of tree stems occurs via symplastic transport in ray parenchyma, Plant Physio., 167 (2015): 963-971. |
| [27] | [ O. Pironneau,M. Tabata, Stability and convergence of a Galerkin-characteristics finite element scheme of lumped mass type, Int. J. Num. Meth. Fluids, 64 (2000): 1240-1253. |
| [28] | [ G. E. Phillips,J. Bodig,J. Goodman, Flow grain analogy, Wood Sci., 14 (1981): 55-64. |
| [29] | [ R. J. Phillips,S. R. Dungan, Asymptotic analysis of flow in sieve tubes with semi-permeable walls, J. Theor. Biol., 162 (1993): 465-485. |
| [30] | [ D. Rotsch, T. Brossard, S. Bihmidine, W. Ying, V. Gaddam, M. Harmata, J. D. Robertson, M. Swyers, S. S. Jurisson and D. M. Braun, Radiosynthesis of 6'-Deoxy-6'[18F]Fluorosucrose via automated synthesis and its utility to study in vivo sucrose transport in maize (Zea mays) leaves PLoS ONE 10 (2015), e0128989. |
| [31] | [ D. Sellier,J. J. Harrington, Phloem transport in trees: A generic surface model, Eco. Mod., 290 (2014): 102-109. |
| [32] | [ D. Sellier,M. J. Plank,J. J. Harrington, A mathematical framework for modelling cambial surface evolution using a level set method, Annals Bot., 108 (2011): 1001-1011. |
| [33] | [ R. Spicer, Symplasmic networks in secondary vascular tissues: Parenchyma distribution and activity supporting long-distance transport, J. Exp. Bot., 65 (2014): 1829-1848. |
| [34] | [ J. F. Swindells, C. F. Snyder, R. C. Hardy and P. E. Golden, Viscosities of sucrose solutions at various temperatures: Tables of recalculated values, NBS Circular 440 (1958). |
| [35] | [ M. V. Thompson,N. M. Holbrook, Application of a single-solute non-steady-state phloem model to the study of long-distance assimilate transport, J. Theo. Bio., 220 (2003): 419-455. |

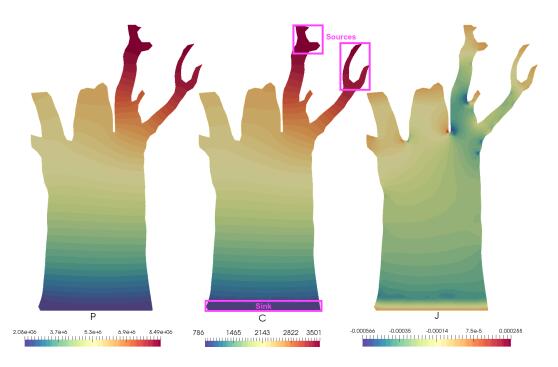
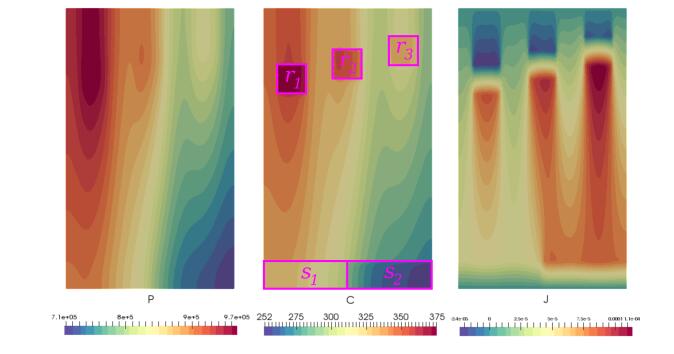
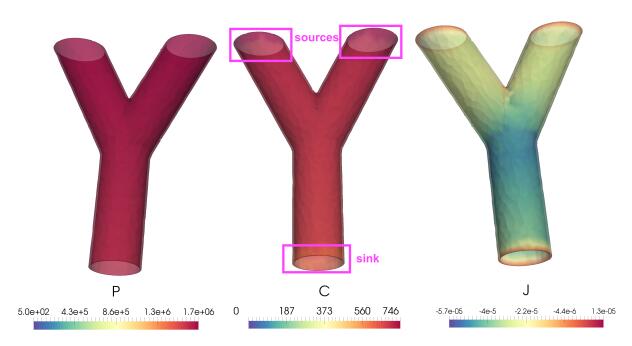
Figures(8) / Tables(2)

Youcef Mammeri, Damien Sellier. A surface model of nonlinear, non-steady-state phloem transport[J]. Mathematical Biosciences and Engineering, 2017, 14(4): 1055-1069. doi: 10.3934/mbe.2017055







 DownLoad:
DownLoad: