Citation: Alacia M. Voth, John G. Alford, Edward W. Swim. Mathematical modeling of continuous and intermittent androgen suppression for the treatment of advanced prostate cancer[J]. Mathematical Biosciences and Engineering, 2017, 14(3): 777-804. doi: 10.3934/mbe.2017043

| [1] | [ P.-A. Abrahamsson, Potential benefits of intermittent androgen suppression therapy in the treatment of prostate cancer: A systematic review of the literature, European Urology, 57 (2010): 49-59. |
| [2] | [ H. T. Banks,S. Dediu,S. L. Ernstberger, Sensitivity functions and their uses in inverse problems, J. Inverse Ill-Posed Probl., 15 (2007): 683-708. |
| [3] | [ H.T. Banks,D.M. Bortz, A parameter sensitivity methodology in the context of HIV delay equation models, J. Math. Biol., 50 (2005): 607-625. |
| [4] | [ N. C. Buchan,S. L. Goldenberg, Intermittent androgen suppression for prostate cancer, Nature Reviews Urology, 7 (2010): 552-560. |
| [5] | [ N. Chitnis,J.M. Hyman,J.M. Chushing, Determining important parameters in the spread of malaria through the sensitivity analysis of a mathematical model, Bull. Math. Biol., 70 (2008): 1272-1296. |
| [6] | [ R.A. Everett,A.M. Packer,Y. Kuang, Can mathematical models predict the outcomes of prostate cancer patients undergoing intermittent androgen deprivation therapy?, Biophys. Rev. Lett., 9 (2014): 139-157. |
| [7] | [ J. K. Hale, Ordinary Differential Equations, 2nd edition, Krieger Publishing, Malabar FL, 1980. |
| [8] | [ Y. Hirata,N. Bruchovsky,K. Aihara, Development of a mathematical model that predicts the outcome of hormone therapy for prostate cancer, J. Theor. Biol., 264 (2010): 517-527. |
| [9] | [ A.M. Ideta,G. Tanaka,T. Takeuchi,K. Aihara, A mathematical model of intermittent androgen suppression for prostate cancer, J. Nonlinear Sci., 18 (2008): 593-614. |
| [10] | [ H. Lepor,N.D. Shore, LHRH agonists for the treatment of prostate cancer: 2012, Reviews in Urology, 14 (2012): 1-12. |
| [11] | [ Prostate cancer treatment (PDQ) -Patient Version National Cancer Institute, 2016. Available from: https://www.cancer.gov/types/prostate/patient/prostate-treatment-pdq. |
| [12] | [ T. Portz,Y. Kuang,J.D. Nagy, A clinical data validated mathematical model of prostate cancer growth under intermittent androgen suppression therapy, AIP Advances, 2 (2012): 1-14. |
| [13] | [ M.H. Rashid,U.B. Chaudhary, Intermittent androgen deprivation therapy for prostate cancer, The Oncologist, 9 (2004): 295-301. |
| [14] | [ F.G. Rick,A.V. Schally, Bench-to-bedside development of agonists and antagonists of luteinizing hormone-releasing hormone for treatment of advanced prostate cancer, Urologic Oncology: Seminars and Original Investigations, 33 (2015): 270-274. |
| [15] | [ A. Sciarra,P.A. Abrahamsson,M. Brausi,M. Galsky,N. Mottet,O. Sartor,T.L.J. Tammela,F.C. da Silva, Intermittent androgen-depravation therapy in prostate cancer: a critical review focused on phase 3 trials, European Urology, 64 (2013): 722-730. |
| [16] | [ L.G. Stanley, Sensitivity equation methods for parameter dependent elliptic equations, Numer. Funct. Anal. Optim., 22 (2001): 721-748. |
| [17] | [ Y. Suzuki,D. Sakai,T. Nomura,Y. Hirata,K. Aihara, A new protocol for intermittent androgen suppresion therapy of prostate cancer with unstable saddle-point dynamics, J. Theor. Biol., 350 (2014): 1-16. |
| [18] | [ G. Tanaka,K. Tsumoto,S. Tsuji,K. Aihara, Analysis on a hybrid systems model of intermittent hormonal therapy for prostate cancer, Physica D, 237 (2008): 2616-2627. |
| [19] | [ F. Verhulst, Nonlinear Differential Equations and Dynamical Systems, 2nd edition, Springer-Verlag, Berlin, 1996. |
| [20] | [ L. Voth, The Exploration and Computations of Mathematical Models of Intermittent Treatment for Prostate Cancer, M. S. thesis, Sam Houston University, 2012. |
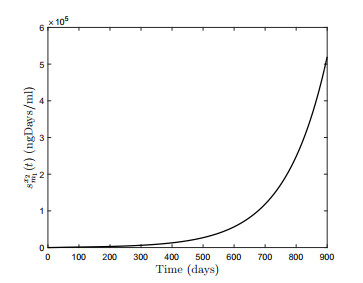
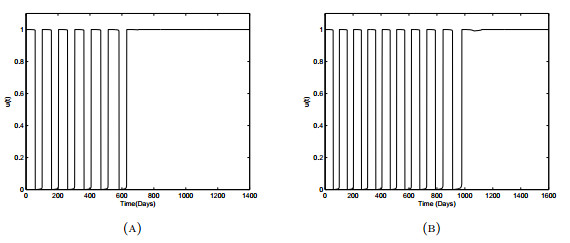
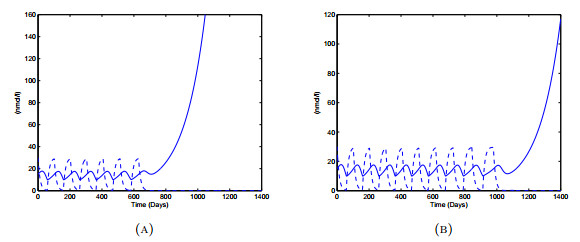
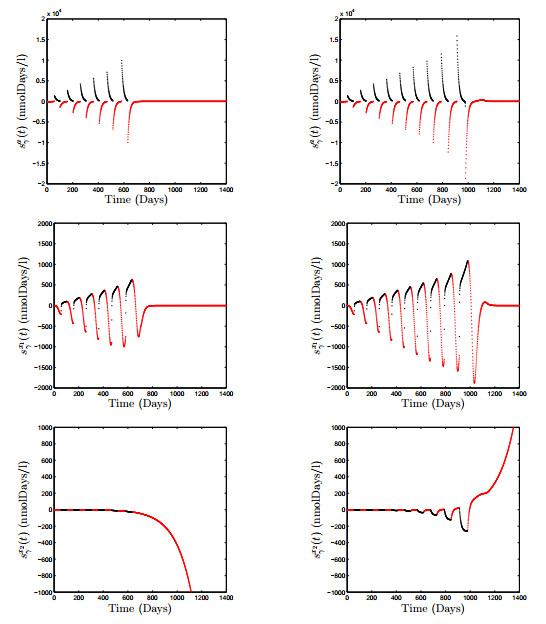
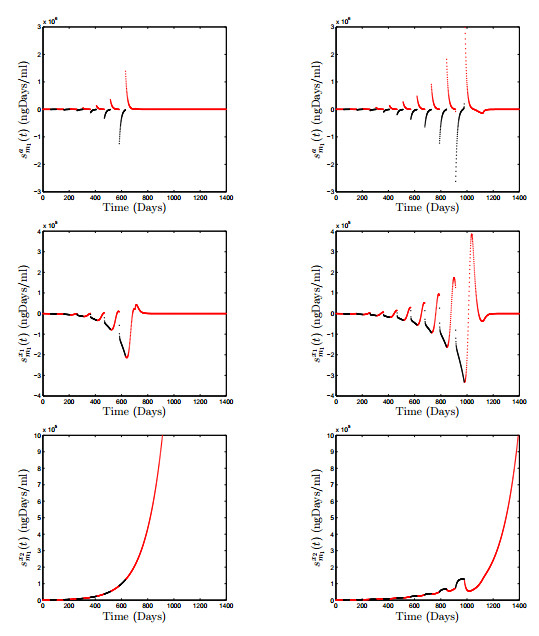
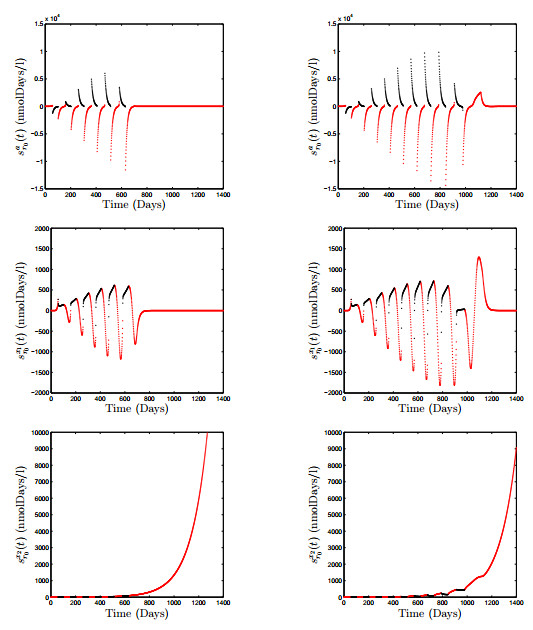
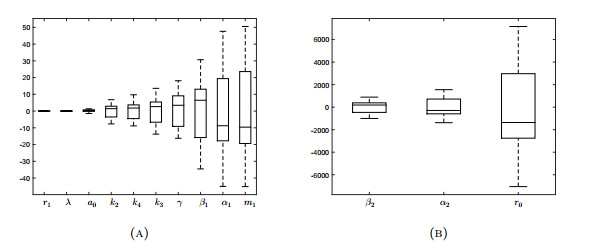
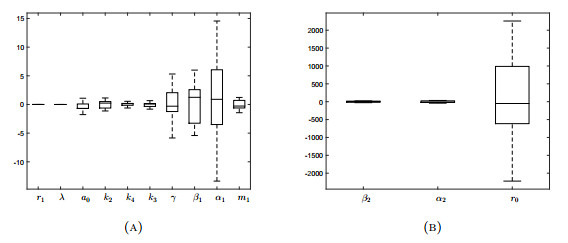
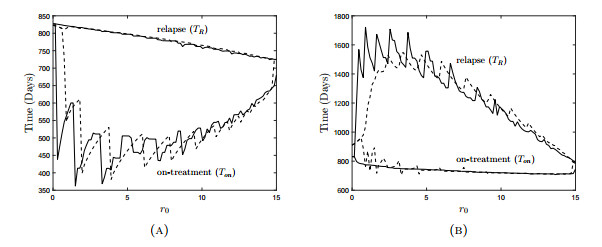
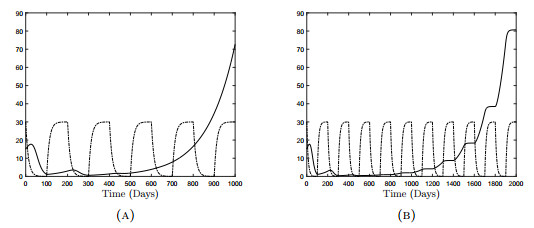
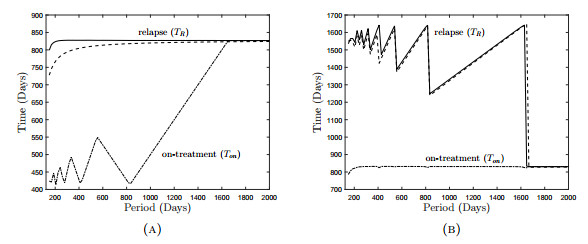
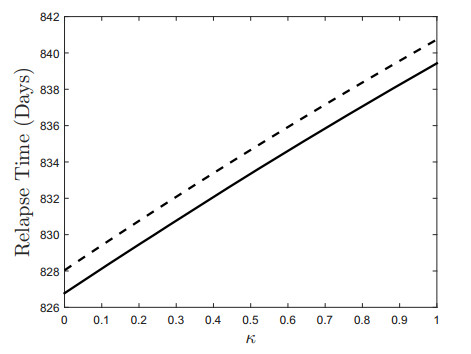
Figures(18) / Tables(3)

Alacia M. Voth, John G. Alford, Edward W. Swim. Mathematical modeling of continuous and intermittent androgen suppression for the treatment of advanced prostate cancer[J]. Mathematical Biosciences and Engineering, 2017, 14(3): 777-804. doi: 10.3934/mbe.2017043







 DownLoad:
DownLoad: