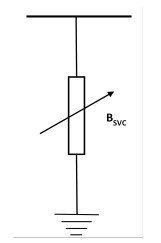
Transmission congestions are caused by electricity trading between generators and distribution companies in a deregulated environment. Power system operation and security in liberalized scenarios are maintained by removing branch overloads. A flexible alternating current transmission system (FACTS) controller is installed in a suitable location to redistribute the power flow among the transmission lines so that the power flows are brought within the capacity of the lines. In this work, series-connected thyristor-controlled switched compensators (TCSCs) and shunt-connected Volt-Ampere reactive (VAR) static compensators (SVCs) are installed in appropriate locations to alter the power flow patterns and to remove overloads. It is proposed to reduce the overload of transmission lines by locating series and shunt connected FACTS devices at proper locations. The size and location of TCSC and SVC devices greatly affect their ability to meet a congestion management goal. An optimization process optimizes the location and size of these devices to maximize the congestion mitigation benefits of the TCSC and SVC controllers. In this work, the whale optimization algorithm (WOA) is used to optimize the value of the objective function by appropriately choosing the location and size of the FACTS controllers. This algorithm has a few parameters that are tuned to give the best overall results. A WOA-based method is proposed to optimize the size and location of the FACTS devices and is implemented on the IEEE-30 bus test case. The results were compared and found to be improved with those of other algorithms such as the particle swarm optimization algorithm (PSO) and the firefly algorithm (FFA).
Citation: Vengadesan Alagapuri, Ashok Bakkiyaraj Radhakrishnan, S. Sakthivel Padaiyatchi. Optimization of position and rating of shunt and series connected FACTS devices for transmission congestion management in deregulated power networks[J]. AIMS Electronics and Electrical Engineering, 2024, 8(2): 165-186. doi: 10.3934/electreng.2024007
Transmission congestions are caused by electricity trading between generators and distribution companies in a deregulated environment. Power system operation and security in liberalized scenarios are maintained by removing branch overloads. A flexible alternating current transmission system (FACTS) controller is installed in a suitable location to redistribute the power flow among the transmission lines so that the power flows are brought within the capacity of the lines. In this work, series-connected thyristor-controlled switched compensators (TCSCs) and shunt-connected Volt-Ampere reactive (VAR) static compensators (SVCs) are installed in appropriate locations to alter the power flow patterns and to remove overloads. It is proposed to reduce the overload of transmission lines by locating series and shunt connected FACTS devices at proper locations. The size and location of TCSC and SVC devices greatly affect their ability to meet a congestion management goal. An optimization process optimizes the location and size of these devices to maximize the congestion mitigation benefits of the TCSC and SVC controllers. In this work, the whale optimization algorithm (WOA) is used to optimize the value of the objective function by appropriately choosing the location and size of the FACTS controllers. This algorithm has a few parameters that are tuned to give the best overall results. A WOA-based method is proposed to optimize the size and location of the FACTS devices and is implemented on the IEEE-30 bus test case. The results were compared and found to be improved with those of other algorithms such as the particle swarm optimization algorithm (PSO) and the firefly algorithm (FFA).

| [1] |
Fang RS, David AK (1999) Optimal Dispatch under transmission contracts. IEEE T Power Syst 14: 732–737. https://doi.org/10.1109/59.761905 doi: 10.1109/59.761905

|
| [2] |
Choudekar P, Asija D (2021) Transmission congestion management using multi objective hybrid flower pollination and particle swarm optimization algorithm by optimal placement of TCSC. Metaheuristic and Evolutionary Computation: Algorithms and Applications, 569–583. https://doi.org/10.1007/978-981-15-7571-6_26 doi: 10.1007/978-981-15-7571-6_26
|
| [3] |
Patel NK, Suthar BN, Thakkar J (2021) Transmission congestion management considering voltage stability margin. SN Applied Sciences 3: 261. https://doi.org/10.1007/s42452-020-04132-9 doi: 10.1007/s42452-020-04132-9
|
| [4] | Gad M, Shinde P, Kulkarni SU (2012) Optimal location of TCSC by sensitivity methods. Int J Comput Eng Res 2: 162–168. |
| [5] |
Leung HC, Chung TS (2000) Optimal power flow with a versatile FACTS controller by genetic algorithm approach. Proc. Power engineering society winter meeting 4: 2806–2811. https://doi.org/10.1049/cp:20000387 doi: 10.1049/cp:20000387
|
| [6] | Hingorani NG, Gyugyi L (1998) Understanding FACTS. The Institution of Electrical and Electronics Engineers, 1998. |
| [7] |
Gyugyi L, Schauder CD, Williams SL, Rietman TR, Torgerson DR, Edris A (1995) The unified power flow controller: a new approach to power transmission control. IEEE T Power Deliver 10: 1085–1097. https://doi.org/10.1109/61.400878 doi: 10.1109/61.400878
|
| [8] |
Larsen E, Millers N, Nilsson S, Lindgren S (1992) Benefits of GTO-based compensation systems for electric utility applications. IEEE T Power Deliver 7: 2056–2064. https://doi.org/10.1109/61.157009 doi: 10.1109/61.157009
|
| [9] |
Preedavinchit P, Srivastava SC (1998) Optimal reactive power dispatch considering FACTS devices. Electric Power Syst Res 46: 251–257. https://doi.org/10.1016/S0378-7796(98)00075-3 doi: 10.1016/S0378-7796(98)00075-3
|
| [10] | Momoh JA, Zhu JZ (1998) A new approach to optimal power flow with phase shifter. SMC'98 Conference Proceedings. 1998 IEEE International Conference on Systems, Man, and Cybernetics (Cat. No. 98CH36218) 5: 4794–4799. |
| [11] |
Liu JY, Song YH (1999) Comparison studies of unified power flow controller with static var compensators and phase shifters. Electric Machines & Power Systems 27: 237–251. https://doi.org/10.1080/073135699269271 doi: 10.1080/073135699269271
|
| [12] |
Besharat H, Taher SA (2008) Congestion management by determining optimal location of TCSC in deregulated power systems. Int J Elec Power 30: 563–568. http://dx.doi.org/10.1016/j.ijepes.2008.08.007 doi: 10.1016/j.ijepes.2008.08.007
|
| [13] |
Singh SN, David AK (2001) Optimal location of FACTS devices for congestion management. Electr Power Syst Res 58: 71–79. https://doi.org/10.1016/S0378-7796(01)00087-6 doi: 10.1016/S0378-7796(01)00087-6
|
| [14] |
Amjady N, Hakimi M (2012) Dynamic voltage stability constrained congestion management framework for deregulated electricity markets. Energy Convers Manag 58: 66–75. https://doi.org/10.1016/j.enconman.2012.01.006 doi: 10.1016/j.enconman.2012.01.006
|
| [15] | Gerbex S, Cherkaoui R, Germond AJ (2003) Optimal location of FACTS devices to enhance power system security. 2003 IEEE Bologna Power Tech Conference Proceedings 3: 7-pp. IEEE. |
| [16] |
Rashed GI, Sun Y (2011) Optimal placement of Thyristor controlled series compensation for enhancing power system security based on computational intelligence techniques. Procedia Engineering 15: 908–914. https://doi.org/10.1016/j.proeng.2011.08.168 doi: 10.1016/j.proeng.2011.08.168
|
| [17] |
Rahim MA, Musirin I, Abidin IZ, Othman MM (2010) Contingency based congestion management and cost minimization using bee colony optimization technique. PECon2010 - 2010 IEEE International Conference on Power and Energy 891–896. https://doi.org/10.1109/PECON.2010.5697705 doi: 10.1109/PECON.2010.5697705
|
| [18] |
Shrestha GB, Feng W (2005) Effects of series compensation on spot price power markets. Int J Elec Power 27: 428–436. https://doi.org/10.1016/j.ijepes.2005.03.001 doi: 10.1016/j.ijepes.2005.03.001
|
| [19] |
Hosseinipoor N, Nabavi SMH (2010) Optimal locating and sizing of TCSC using genetic Algorithm for congestion management in deregualted power markets. 2010 9th International Conference on Environment and Electrical Engineering 136–139. https://doi.org/10.1109/EEEIC.2010.5489968 doi: 10.1109/EEEIC.2010.5489968
|
| [20] |
Biswas PP, Arora P, Mallipeddi R, Suganthan PN, Panigrahi BK (2021) Optimal placement and sizing of FACTS devices for optimal power flow in a wind power integrated electrical network. Neural Comput Appl 33: 6753–6774. https://doi.org/10.1007/s00521-020-05453-x doi: 10.1007/s00521-020-05453-x
|
| [21] | Verma KS, Singh SN, Gupta HO (2001) FACTS devices location for enhancement of total transfer capability. 2001 IEEE Power Engineering Society Winter Meeting. Conference Proceedings (Cat. No. 01CH37194) 2: 522–527. |
| [22] |
Jordehi AR (2015) Brainstorm optimisation algorithm (BSOA): An efficient algorithm for finding optimal location and setting of FACTS devices in electric power systems. Int J Elec Power 69: 48–57. https://doi.org/10.1016/j.ijepes.2014.12.083 doi: 10.1016/j.ijepes.2014.12.083
|
| [23] |
Mirjalili S, Lewis A (2016) The whale optimization algorithm. Adv Eng Softw 95: 51-67. https://doi.org/10.1016/j.advengsoft.2016.01.008 doi: 10.1016/j.advengsoft.2016.01.008
|
| [24] | The IEEE 30-Bus Test System. Available from: http://www.ee.washington.edu/research/pstca/pf30/pg$_$tca30bus.htm. |






Figures(7) / Tables(16)

Vengadesan Alagapuri, Ashok Bakkiyaraj Radhakrishnan, S. Sakthivel Padaiyatchi. Optimization of position and rating of shunt and series connected FACTS devices for transmission congestion management in deregulated power networks[J]. AIMS Electronics and Electrical Engineering, 2024, 8(2): 165-186. doi: 10.3934/electreng.2024007







 DownLoad:
DownLoad: