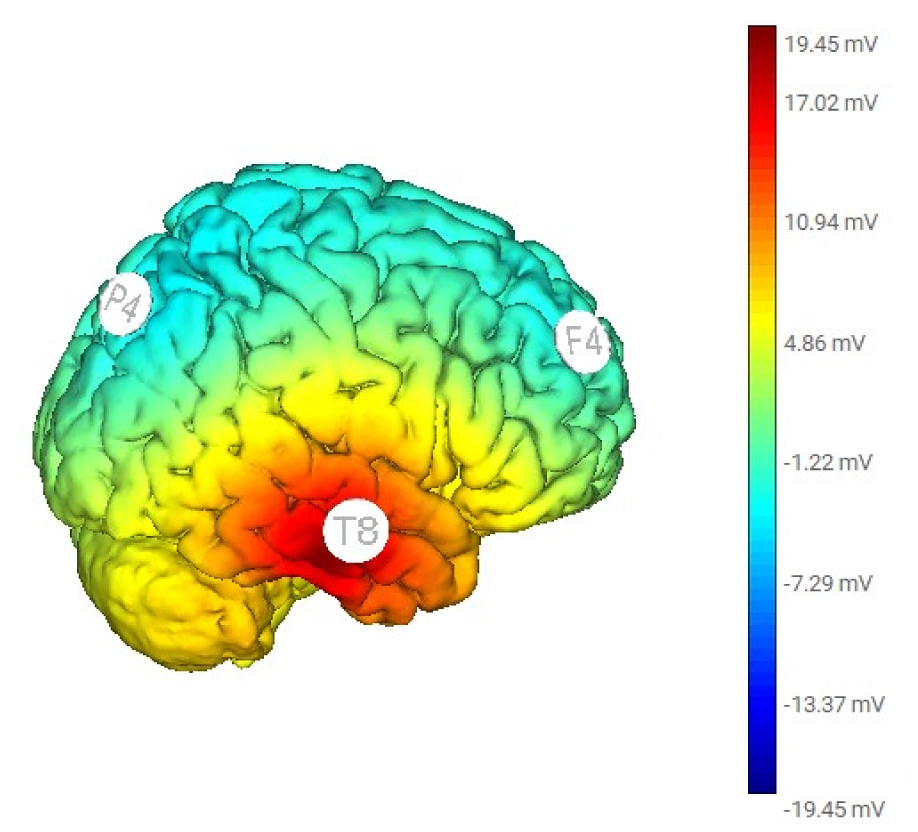
Transcranial alternating current stimulation (tACS) at 5-Hz to the right hemisphere can alleviate anxiety symptoms. We aimed to explore the connectivity changes following the treatment. We collected electroencephalography (EEG) data from 24 participants with anxiety disorders before and after the tACS treatment during a single session. Electric stimulation was applied over the right hemisphere, with 1.0 mA at F4, 1.0 mA at P4, and 2.0 mA at T8, following the 10-10 EEG convention. With eLORETA, the scalp signals were transformed into the cortex's current source density. We assessed the connectivity changes at theta frequency between the centers of Brodmann area (BA) 6/8 (frontal), BA 39/40 (parietal), and BA 21 (middle temporal). Functional connectivity was indicated by lagged coherences and lagged phase synchronization. Paired t-tests were used to quantify the differences statistically. We observed enhanced lagged phase synchronization at theta frequency between the frontal and parietal regions (P = 0.002) and between the parietal and temporal regions (P = 0.005) after Bonferroni correction. Applying tACS 5-Hz over the right hemisphere enhanced inter-regional interaction, which was spectrum-specific and mainly mediated by phase rather than power synchrony. The potential neural mechanisms are discussed.
Citation: Tien-Wen Lee, Gerald Tramontano. Connectivity changes following transcranial alternating current stimulation at 5-Hz: an EEG study[J]. AIMS Neuroscience, 2024, 11(4): 439-448. doi: 10.3934/Neuroscience.2024026
Transcranial alternating current stimulation (tACS) at 5-Hz to the right hemisphere can alleviate anxiety symptoms. We aimed to explore the connectivity changes following the treatment. We collected electroencephalography (EEG) data from 24 participants with anxiety disorders before and after the tACS treatment during a single session. Electric stimulation was applied over the right hemisphere, with 1.0 mA at F4, 1.0 mA at P4, and 2.0 mA at T8, following the 10-10 EEG convention. With eLORETA, the scalp signals were transformed into the cortex's current source density. We assessed the connectivity changes at theta frequency between the centers of Brodmann area (BA) 6/8 (frontal), BA 39/40 (parietal), and BA 21 (middle temporal). Functional connectivity was indicated by lagged coherences and lagged phase synchronization. Paired t-tests were used to quantify the differences statistically. We observed enhanced lagged phase synchronization at theta frequency between the frontal and parietal regions (P = 0.002) and between the parietal and temporal regions (P = 0.005) after Bonferroni correction. Applying tACS 5-Hz over the right hemisphere enhanced inter-regional interaction, which was spectrum-specific and mainly mediated by phase rather than power synchrony. The potential neural mechanisms are discussed.

| [1] |
Antal A, Boros K, Poreisz C, et al. (2008) Comparatively weak after-effects of transcranial alternating current stimulation (tACS) on cortical excitability in humans. Brain Stimul 1: 97-105. https://doi.org/10.1016/j.brs.2007.10.001 
|
| [2] |
Bland NS, Sale MV (2019) Current challenges: the ups and downs of tACS. Exp Brain Res 237: 3071-3088. https://doi.org/10.1007/s00221-019-05666-0
|
| [3] |
Clancy KJ, Baisley SK, Albizu A, et al. (2018) Transcranial alternating current stimulation induces long-term augmentation of neural connectivity and sustained anxiety reduction. Soc Cogn Affect Neur 13: 1305-1316. https://doi.org/10.1093/scan/nsy096
|
| [4] |
Lee TW, Li CS R, Tramontano G (2024) Tripod transcranial alternating current stimulation at 5-Hz over right hemisphere may relieve anxiety symptoms: a preliminary report. J Affect Disord 360: 156-162. https://doi.org/10.1016/j.jad.2024.05.166
|
| [5] | Lee TW, Tramontano G (2024) Neural consequences of 5-Hz transcranial alternating current stimulation over right hemisphere: an eLORETA EEG study. Neurosci Lett 835. https://doi.org/10.1016/j.neulet.2024.137849 |
| [6] |
Engel K, Bandelow B, Gruber O, et al. (2009) Neuroimaging in anxiety disorders. J Neural Transm (Vienna) 116: 703-716. https://doi.org/10.1007/s00702-008-0077-9
|
| [7] |
Spitzer B, Haegens S (2017) Beyond the status quo: a role for beta oscillations in endogenous content (re) activation. eNeuro 4. https://doi.org/10.1523/ENEURO.0170-17.2017
|
| [8] |
Oathes DJ, Ray WJ, Yamasaki AS, et al. (2008) Worry, generalized anxiety disorder, and emotion: Evidence from the eeg gamma band. Biol Psychol 79: 165-170. https://doi.org/10.1016/j.biopsycho.2008.04.005
|
| [9] |
Miskovic V, Ashbaugh AR, Santesso DL, et al. (2010) Frontal brain oscillations and social anxiety: a cross-frequency spectral analysis during baseline and speech anticipation. Biol Psychol 83: 125-132. https://doi.org/10.1016/j.biopsycho.2009.11.010
|
| [10] |
Berger H (1929) Ueber das elektroenkephalogramm des menschen. Archiv für Psychiatrie und Nervenkrankheiten 87: 527-570. https://doi.org/10.1007/BF01797193
|
| [11] | Sharma A, Singh M (2015) Assessing alpha activity in attention and relaxed state: An EEG analysis. IEEE : 508-513. https://doi.org/10.1109/NGCT.2015.7375171 |
| [12] |
Helfrich RF, Schneider TR, Rach S, et al. (2014) Entrainment of brain oscillations by transcranial alternating current stimulation. Curr Biol 24: 333-339. https://doi.org/10.1016/j.cub.2013.12.041
|
| [13] |
Voss U, Holzmann R, Hobson A, et al. (2014) Induction of self awareness in dreams through frontal low current stimulation of gamma activity. Nat Neurosci 17: 810-812. https://doi.org/10.1038/nn.3719
|
| [14] |
Shekelle PG, Cook IA, Miake-Lye IM, et al. (2018) The effectiveness and risks of cranial electrical stimulation for the treatment of pain, depression, anxiety, PTSD, and insomnia: A systematic review. Ann Intern Med 168: 414-421. https://doi.org/10.7326/M17-1970
|
| [15] |
Anastassiou CA, Montgomery SM, Barahona M, et al. (2010) The effect of spatially inhomogeneous extracellular electric fields on neurons. J Neurosci 30: 1925-1936. https://doi.org/10.1523/JNEUROSCI.3635-09.2010
|
| [16] |
Radman T, Su Y, An JH, et al. (2007) Spike timing amplifies the effect of electric fields on neurons: implications for endogenous field effects. J Neurosci 27: 3030-3036. https://doi.org/10.1523/JNEUROSCI.0095-07.2007
|
| [17] |
Radman T, Ramos RL, Brumberg JC, et al. (2009) Role of cortical cell type and morphology in subthreshold and suprathreshold uniform electric field stimulation in vitro. Brain Stimul 2: 215-228, 228 e211-213. https://doi.org/10.1016/j.brs.2009.03.007
|
| [18] |
Krause MR, Vieira PG, Csorba BA, et al. (2019) Transcranial alternating current stimulation entrains single-neuron activity in the primate brain. Proc Natl Acad Sci U S A 116: 5747-5755. https://doi.org/10.1073/pnas.1815958116
|
| [19] |
Levy W, Steward O (1983) Temporal contiguity requirements for long-term associative potentiation/depression in the hippocampus. Neuroscience 8: 791-797. https://doi.org/10.1016/0306-4522(83)90010-6
|
| [20] | Markram H, Gerstner W, Sjöström PJ (2012) Spike-timing-dependent plasticity: a comprehensive overview. Front Synaptic Neuro 4: 2. https://doi.org/10.3389/fnsyn.2012.00002 |
| [21] | Pascual-Marqui RD (2007) Discrete, 3D distributed, linear imaging methods of electric neuronal activity. Part 1: exact, zero error localization. arXiv preprint : arXiv:07103341. |
| [22] |
Jurcak V, Tsuzuki D, Dan I (2007) 10/20, 10/10, and 10/5 systems revisited: their validity as relative head-surface-based positioning systems. Neuroimage 34: 1600-1611. https://doi.org/10.1016/j.neuroimage.2006.09.024
|
| [23] | Pascual-Marqui RD (2007) Instantaneous and lagged measurements of linear and nonlinear dependence between groups of multivariate time series: frequency decomposition. arXiv preprint : arXiv:07111455. |
| [24] |
Beck AT, Epstein N, Brown G, et al. (1993) Beck anxiety inventory. J Consult Clin Psych 56: 893-897. https://doi.org/10.1037/0022-006X.56.6.893
|
| [25] |
Delorme A, Makeig S (2004) EEGLAB: an open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. J Neurosci Methods 134: 9-21. https://doi.org/10.1016/j.jneumeth.2003.10.009
|
| [26] |
Vossen A, Gross J, Thut G (2015) Alpha Power Increase After Transcranial Alternating Current Stimulation at Alpha Frequency (alpha-tACS) Reflects Plastic Changes Rather Than Entrainment. Brain Stimul 8: 499-508. https://doi.org/10.1016/j.brs.2014.12.004
|
| [27] |
Lee TW, Xue SW (2018) Functional connectivity maps based on hippocampal and thalamic dynamics may account for the default-mode network. Eur J Neurosci 47: 388-398. https://doi.org/10.1111/ejn.13828
|
| [28] |
Jones KT, Johnson EL, Gazzaley A, et al. (2022) Structural and functional network mechanisms of rescuing cognitive control in aging. Neuroimage 262: 119547. https://doi.org/10.1016/j.neuroimage.2022.119547
|
| [29] |
Davis MC, Fitzgerald PB, Bailey NW, et al. (2023) Effects of medial prefrontal transcranial alternating current stimulation on neural activity and connectivity in people with Huntington's disease and neurotypical controls. Brain Res 1811: 148379. https://doi.org/10.1016/j.brainres.2023.148379
|
| [30] |
Aktürk T, de Graaf TA, Güntekin B, et al. (2022) Enhancing memory capacity by experimentally slowing theta frequency oscillations using combined EEG-tACS. Sci Rep 12: 14199. https://doi.org/10.1038/s41598-022-18665-z
|
| [31] | Yeh TC, Huang CC, Chung YA, et al. (2023) Online Left-Hemispheric In-Phase Frontoparietal Theta tACS Modulates Theta-Band EEG Source-Based Large-Scale Functional Network Connectivity in Patients with Schizophrenia: A Randomized, Double-Blind, Sham-Controlled Clinical Trial. Biomedicines 11. https://doi.org/10.3390/biomedicines11020630 |
| [32] |
Lehmann D, Faber PL, Tei S, et al. (2012) Reduced functional connectivity between cortical sources in five meditation traditions detected with lagged coherence using EEG tomography. Neuroimage 60: 1574-1586. https://doi.org/10.1016/j.neuroimage.2012.01.042
|
| [33] | Pascual-Marqui R, Biscay R, Bosch-Bayard J, et al. (2014) Isolated effective coherence (iCoh): causal information flow excluding indirect paths. arXiv preprint : arXiv:14024887. |
| [34] |
Brignani D, Ruzzoli M, Mauri P, et al. (2013) Is transcranial alternating current stimulation effective in modulating brain oscillations?. PLoS One 8: e56589. https://doi.org/10.1371/journal.pone.0056589
|
| [35] |
Alexander ML, Alagapan S, Lugo CE, et al. (2019) Double-blind, randomized pilot clinical trial targeting alpha oscillations with transcranial alternating current stimulation (tACS) for the treatment of major depressive disorder (MDD). Transl Psychiatry 9: 106. https://doi.org/10.1038/s41398-019-0439-0
|
| [36] |
Lafon B, Henin S, Huang Y, et al. (2017) Low frequency transcranial electrical stimulation does not entrain sleep rhythms measured by human intracranial recordings. Nat Commun 8: 1199. https://doi.org/10.1038/s41467-017-01045-x
|
| [37] |
Ross ED (2021) Differential hemispheric lateralization of emotions and related display behaviors: emotion-type hypothesis. Brain Sci 11: 1034. https://doi.org/10.3390/brainsci11081034
|

Figures(2) / Tables(1)

Tien-Wen Lee, Gerald Tramontano. Connectivity changes following transcranial alternating current stimulation at 5-Hz: an EEG study[J]. AIMS Neuroscience, 2024, 11(4): 439-448. doi: 10.3934/Neuroscience.2024026







 DownLoad:
DownLoad: