Citation: Rinaldo M. Colombo, Elena Rossi. A modeling framework for biological pest control[J]. Mathematical Biosciences and Engineering, 2020, 17(2): 1413-1427. doi: 10.3934/mbe.2020072

| [1] | K. Wickwire, Mathematical models for the control of pests and infectious diseases: a survey, Theor. Popul. Biol., 11 (1977), 182-238. |
| [2] | J. Grasman, O. A. van Herwaarden, L. Hemerik, J. C. van Lenteren, A two-component model of host-parasitoid interactions: determination of the size of inundative releases of parasitoids in biological pest control, Math. Biosci., 169 (2001), 207-216. |
| [3] | G. E. Heimpel, N. J. Mills, Biological control: Ecology and applications, Biological Control: Ecology and Applications, Cambridge University Press, 2017. Available from: https://doi.org/10.1017/9781139029117. |
| [4] | J. Jiao, S. Cai, L. Li, Y. Zhang, Dynamics of a predator-prey model with impulsive biological control and unilaterally impulsive diffusion, Adv. Differ. Equ., 2016 (2016), 150. Available from: https://doi.org/10.1186/s13662-016-0851-1. |
| [5] | M. Rafikov, J. M. Balthazar, H. F. Von Bremen, Management of complex systems: Modeling the biological pest control, Biophys. Rev. Lett., 03 (2008), 241-256. |
| [6] | C. M. Dafermos, Hyperbolic conservation laws in continuum physics, vol. 325 of Grundlehren der Mathematischen Wissenschaften [Fundamental Principles of Mathematical Sciences], 4th edition, Springer-Verlag, Berlin, 2016. Available from: http://dx.doi.org/10.1007/ 978-3-662-49451-6. |
| [7] | J. D. Murray, Mathematical biology. I An introduction, vol. 17 of Interdisciplinary Applied Mathematics, 3rd edition, Springer-Verlag, New York, 2002. Available from: https://doi.org/10.1007/b98868. |
| [8] | R. M. Colombo, E. Rossi, Hyperbolic predators vs. parabolic prey, Commun. Math. Sci., 13 (2015), 369-400. |
| [9] | R. M. Colombo, E. Rossi, Control in a mixed hyperbolic-parabolic predator prey model, 2019, Preprint. |
| [10] | M. Colombo, G. Crippa, L. V. Spinolo, On the singular local limit for conservation laws with nonlocal fluxes, Arch. Ration. Mech. Anal., 233 (2019), 1131-1167. |
| [11] | R. J. LeVeque, Finite volume methods for hyperbolic problems, Cambridge Texts in Applied Mathematics, Cambridge University Press, Cambridge, 2002. Available from: http://dx.doi.org/10.1017/CBO9780511791253. |
| [12] | E. Rossi, V. Schleper, Convergence of numerical scheme for a mixed hyperbolic-parabolic system in two space dimensions, ESAIM Math. Model. Numer. Anal., 50 (2016), 475-497. |
| [13] | R. Bürger, G. Chowell, E. Gavilán, P. Mulet, L. M. Villada, Numerical solution of a spatio-temporal gender-structured model for hantavirus infection in rodents, Math. Biosci. Eng., 15 (2018), 95-123. |
| [14] | R. Bürger, G. Chowell, E. Gavilán, P. Mulet, L. M. Villada, Numerical solution of a spatio-temporal predator-prey model with infected prey, Math. Biosci. Eng., 16 (2019), 438-473. |
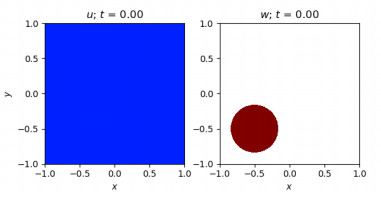
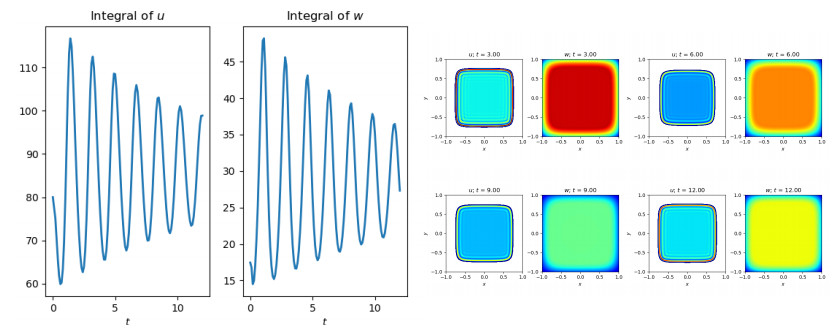
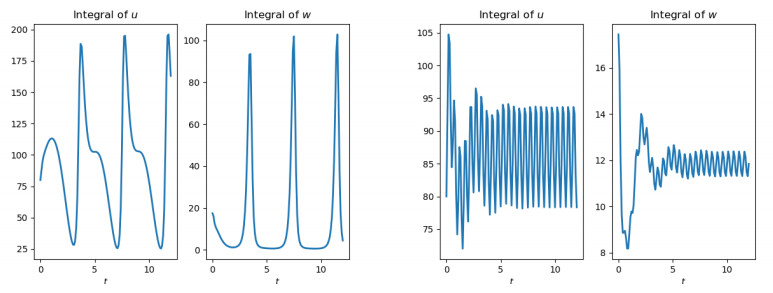
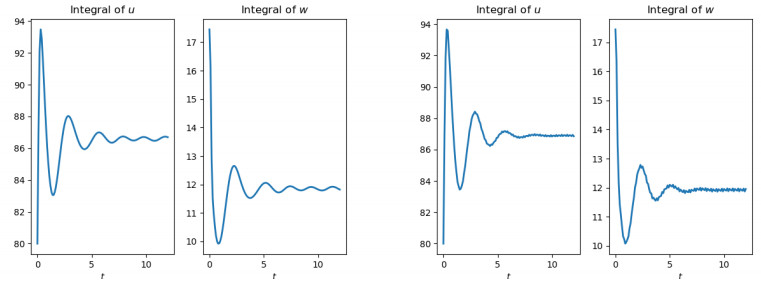
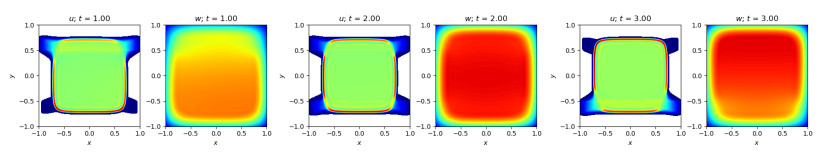
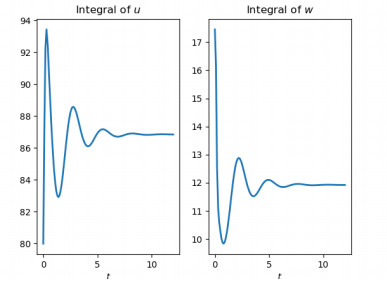
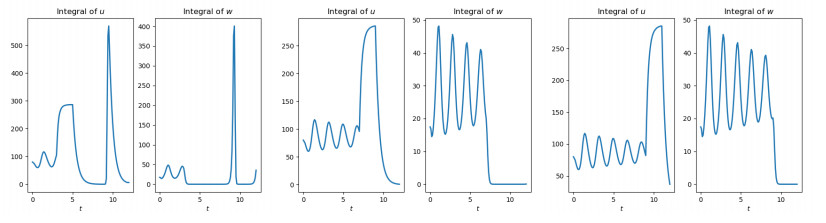
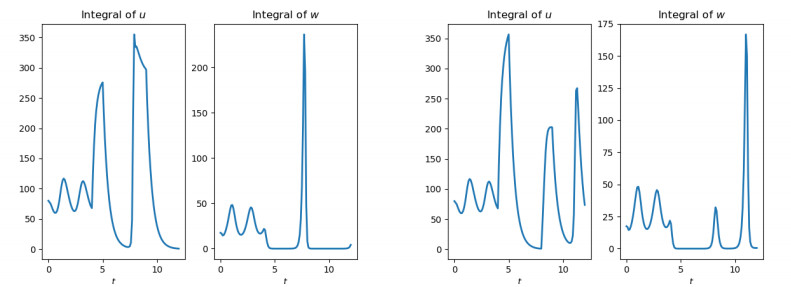
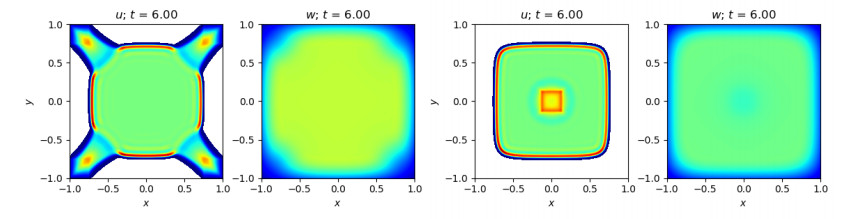
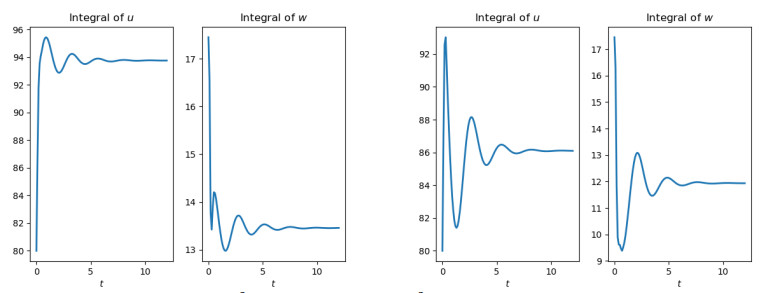
Figures(10)

Rinaldo M. Colombo, Elena Rossi. A modeling framework for biological pest control[J]. Mathematical Biosciences and Engineering, 2020, 17(2): 1413-1427. doi: 10.3934/mbe.2020072







 DownLoad:
DownLoad: