Citation: George A. Michael, Raphaël Mizzi, Cyril Couffe, Germán Gálvez-García, Élodie Labeye. Spotting from The Rightmost Deep: A Temporal Field Advantage in A Behavioural Task of Attention And Filtering[J]. AIMS Neuroscience, 2016, 3(1): 56-66. doi: 10.3934/Neuroscience.2016.1.56

| [1] |
Donnelly N, Humphreys GW, Riddoch MJ (1991) Parallel computation of primitive shape descriptions. J Exp Psychol Hum Percept Perform 17: 561-570. doi: 10.1037/0096-1523.17.2.561

|
| [2] | Krauzlis RJ, Lovejoy LP, Zenon A (2013) Superior Colliculus and Visual Spatial Attention. Annu Rev Neurosci 36. doi:10.1146/annurev-neuro-062012-170249. |
| [3] | Wurtz RH, McAlonan K, Cavanaugh J, et al. (2011) Thalamic pathways for active vision. Trends Cogn Sci 15: 177-184. doi:10.1016/j.tics.2011.02.004. |
| [4] | Fischer J, Whitney D (2012) Attention gates visual coding in the human pulvinar. Nat Commun 3: 1051. doi:10.1038/ncomms2054. |
| [5] |
Strumpf H, Mangun GR, Boehler CN, et al. (2013) The role of the pulvinar in distractor processing and visual search. Hum Brain Mapp 34: 1115-1132. doi:10.1002/hbm.21496. doi: 10.1002/hbm.21496
|
| [6] |
Michael GA, Desmedt S (2004) The human pulvinar and attentional processing of visual distractors. Neurosci Lett 362: 176-181. doi:10.1016/j.neulet.2004.01.062. doi: 10.1016/j.neulet.2004.01.062
|
| [7] | Snow JC, Allen HA, Rafal RD, et al. (2009) Impaired attentional selection following lesions to human pulvinar: Evidence for homology between human and monkey. Proc Natl Acad Sci USA 106: 4054-4059. doi:10.1073/pnas.0810086106. |
| [8] |
Lyon DC, Nassi JJ, Callaway EM (2010) A Disynaptic Relay from Superior Colliculus to Dorsal Stream Visual Cortex in Macaque Monkey. Neuron 65: 270-279. doi:10.1016/j.neuron.2010.01.003. doi: 10.1016/j.neuron.2010.01.003
|
| [9] | Goldberg ME, Wurtz RH (1972) Activity of superior colliculus in behaving monkey. I. Visual receptive fields of single neurons. J Neurophysiol 35: 542-559. |
| [10] |
Itaya SK, Van Hoesen GW (1983) Retinal projections to the inferior and medial pulvinar nuclei in the old-world monkey. Brain Res 269: 223-230. doi:10.1016/0006-8993(83)90131-2. doi: 10.1016/0006-8993(83)90131-2
|
| [11] |
Wilson ME, Toyne MJ (1970) Retino-tectal and cortico-tectal projections inMacaca mulatta. Brain Res 24: 395-406. doi:10.1016/0006-8993(70)90181-2. doi: 10.1016/0006-8993(70)90181-2
|
| [12] |
Cotton PL, Smith AT (2007) Contralateral Visual Hemifield Representations in the Human Pulvinar Nucleus. J Neurophysiol 98: 1600-1609. doi:10.1152/jn.00419.2007. doi: 10.1152/jn.00419.2007
|
| [13] |
Schneider KA, Kastner S (2005) Visual Responses of the Human Superior Colliculus: A High-Resolution Functional Magnetic Resonance Imaging Study. J Neurophysiol 94: 2491-2503. doi:10.1152/jn.00288.2005. doi: 10.1152/jn.00288.2005
|
| [14] |
Jóhannesson ÓI, Kristjánsson Á (2013) Violating the main sequence: asymmetries in saccadic peak velocities for saccades into the temporal versus nasal hemifields. Exp Brain Res 227: 101-110. doi:10.1007/s00221-013-3490-8. doi: 10.1007/s00221-013-3490-8
|
| [15] |
Rafal R, Henik A, Smith J (1991) Extrageniculate Contributions to Reflex Visual Orienting in Normal Humans: A Temporal Hemifield Advantage. J Cogn Neurosci 3: 322–328. doi:10.1162/jocn.1991.3.4.322. doi: 10.1162/jocn.1991.3.4.322
|
| [16] |
Zackon DH, Casson EJ, Zafar A, et al. (1999) The temporal order judgment paradigm: subcorticalattentional contribution under exogenous and endogenouscueing conditions. Neuropsychologia 37: 511-520. doi:10.1016/S0028-3932(98)00134-1. doi: 10.1016/S0028-3932(98)00134-1
|
| [17] |
Michael GA, Ojéda N (2005) Visual field asymmetries in selective attention: Evidence from a modified search paradigm. Neurosci Lett 388: 65-70. doi:10.1016/j.neulet.2005.06.027. doi: 10.1016/j.neulet.2005.06.027
|
| [18] |
Michael GA, Gálvez-García G (2011) Salience-based progression of visual attention. Behav Brain Res 224: 87-99. doi:10.1016/j.bbr.2011.05.024. doi: 10.1016/j.bbr.2011.05.024
|
| [19] | Rogers LJ, Vallortigara G, Andrew RJ (2013) Divided brains: The biology and behavior of brain asymmetries. Cambridge, UK: Cambridge University Press. |
| [20] |
LaBerge D (1990) Thalamic and Cortical Mechanisms of Attention Suggested by Recent Positron Emission Tomographic Experiments. J Cogn Neurosci 2: 358-372. doi:10.1162/jocn.1990.2.4.358. doi: 10.1162/jocn.1990.2.4.358
|
| [21] |
Oldfield RC (1971) The assessment and analysis of handedness: the Edinburgh inventory. Neuropsychologia 9: 97-113. doi: 10.1016/0028-3932(71)90067-4
|
| [22] | Christman S (1989) Perceptual characteristics in visual laterality research. Brain Cogn 11: 238-257. |
| [23] |
Williams C, Azzopardi P, Cowey A (1995) Nasal and temporal retinal ganglion cells projecting to the midbrain: Implications for “blindsight.” Neuroscience 65: 577-586. doi:10.1016/0306-4522(94)00489-R. doi: 10.1016/0306-4522(94)00489-R
|
| [24] | Sylvester R, Josephs O, Driver J, et al. (2007) Visual fMRI Responses in Human Superior Colliculus Show a Temporal–Nasal Asymmetry That Is Absent in Lateral Geniculate and Visual Cortex. J Neurophysiol 97: 1495-1502. doi:10.1152/jn.00835.2006. |
| [25] | Sapir A, Soroker N, Berger A, et al. (1992) Inhibition of return in spatial attention: direct evidence for collicular generation. Nat Neurosci 2: 1053-1054. doi:10.1038/15977. |
| [26] |
Sapir A, Rafal R, Henik A (2002) Attending to the thalamus: inhibition of return and nasal-temporal asymmetry in the pulvinar. Neuroreport 13: 693-697. doi: 10.1097/00001756-200204160-00031
|
| [27] |
Mesulam MM (1999) Spatial attention and neglect: parietal, frontal and cingulate contributions to the mental representation and attentional targeting of salient extrapersonal events. Philos Trans R Soc Lond B Biol Sci 354: 1325-1346. doi: 10.1098/rstb.1999.0482
|
| [28] |
Karnath H-O, Himmelbach M, Rorden C (2002) The subcortical anatomy of human spatial neglect: putamen, caudate nucleus and pulvinar. Brain 125: 350-360. doi: 10.1093/brain/awf032
|
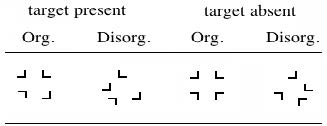
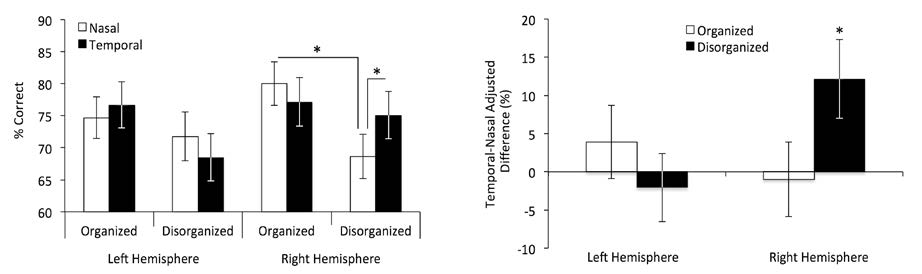
Figures(2) / Tables(1)

George A. Michael, Raphaël Mizzi, Cyril Couffe, Germán Gálvez-García, Élodie Labeye. Spotting from The Rightmost Deep: A Temporal Field Advantage in A Behavioural Task of Attention And Filtering[J]. AIMS Neuroscience, 2016, 3(1): 56-66. doi: 10.3934/Neuroscience.2016.1.56







 DownLoad:
DownLoad: