Citation: Eric Ariel L. Salas, Virginia A. Seamster, Kenneth G. Boykin, Nicole M. Harings, Keith W. Dixon. Modeling the impacts of climate change on Species of Concern (birds) in South Central U.S. based on bioclimatic variables[J]. AIMS Environmental Science, 2017, 4(2): 358-385. doi: 10.3934/environsci.2017.2.358

| [1] |
Huntley B, Collingham YC, Willis SG, et al. (2008) Potential impacts of climatic change on European breeding birds. PLOS ONE 3: e1439. doi: 10.1371/journal.pone.0001439

|
| [2] | Beaumont LJ, Pitman AJ, Poulsen M, et al. (2007) Where will species go? Incorporating new advances in climate modelling into projections of species distributions. Glob Change Biol 13: 1368-1385. |
| [3] |
Marini MA, Barbet-Massin M, Lopes LE, et al. (2009) Predicted climate-driven bird distribution changes and forecasted conservation conflicts in a neotropical savanna. Conserv Biol J Soc Conserv Biol 23: 1558-1567. doi: 10.1111/j.1523-1739.2009.01258.x
|
| [4] | Şekercioğlu ÇH, Primack RB, Wormworth J (2012) The effects of climate change on tropical birds Biol Conserv 148: 1-18. |
| [5] |
Gregory RD (2009) An indicator of the impact of climatic change on European bird populations. PLOS ONE 4: e4678. doi: 10.1371/journal.pone.0004678
|
| [6] |
Julliard R, Jiguet F, Couvet D (2004) Evidence for the impact of global warming on the long–term population dynamics of common birds. Proc R Soc Lond B Biol Sci 271: 490-492. doi: 10.1098/rsbl.2004.0229
|
| [7] |
Berry PM, Dawson TP, Harrison PA, et al. (2002) Modelling potential impacts of climate change on the bioclimatic envelope of species in Britain and Ireland. Glob Ecol Biogeogr 11: 453-462. doi: 10.1111/j.1466-8238.2002.00304.x
|
| [8] |
Thuiller W (2004) Patterns and uncertainties of species' range shifts under climate change. Glob Change Biol 10: 2020-2027. doi: 10.1111/j.1365-2486.2004.00859.x
|
| [9] |
Sax DF (2007) Ecological and evolutionary insights from species invasions. Trends Ecol Evol 22: 465-471. doi: 10.1016/j.tree.2007.06.009
|
| [10] |
Geyer J (2011) Classification of climate-change-induced stresses on biological diversity. Conserv Biol 25: 708-715. doi: 10.1111/j.1523-1739.2011.01676.x
|
| [11] |
Foden WB (2013) Identifying the world's most climate change vulnerable species: A systematic trait-based assessment of all birds, amphibians and corals. PLOS ONE 8: e65427. doi: 10.1371/journal.pone.0065427
|
| [12] |
Jeschke JM, Strayer DL (2008) Usefulness of bioclimatic models for studying climate change and invasive species. Ann N Y Acad Sci 1134: 1-24. doi: 10.1196/annals.1439.002
|
| [13] |
Dawson TP, Jackson ST, House JI, et al. (2011) Beyond predictions: Biodiversity conservation in a changing climate. Science 332: 53-58. doi: 10.1126/science.1200303
|
| [14] |
Rees M, Paull DJ, Carthew SW (2007) Factors influencing the distribution of the yellow-bellied glider (Petaurus australis australis) in Victoria, Australia. Wildl Res 34: 228. doi: 10.1071/WR06027
|
| [15] |
Soberón J (2007) Grinnellian and Eltonian niches and geographic distributions of species. Ecol Lett 10: 1115-1123. doi: 10.1111/j.1461-0248.2007.01107.x
|
| [16] | Peterson AT (2011) Ecological Niches and Geographic Distributions. Princeton, N.J: Princeton University Press. |
| [17] |
Guisan A, Thuiller W (2005) Predicting species distribution: offering more than simple habitat models. Ecol Lett 8: 993-1009. doi: 10.1111/j.1461-0248.2005.00792.x
|
| [18] |
Beale CM, Lennon JJ, Gimona A (2008) Opening the climate envelope reveals no macroscale associations with climate in European birds. Proc Natl Acad Sci 105: 14908-14912. doi: 10.1073/pnas.0803506105
|
| [19] |
Jiménez-Valverde A (2011) Dominant climate influences on North American bird distributions. Glob Ecol Biogeogr 20: 114-118. doi: 10.1111/j.1466-8238.2010.00574.x
|
| [20] |
Pearson RG, Dawson TP (2003) Predicting the impacts of climate change on the distribution of species: are bioclimate envelope models useful? Glob Ecol Biogeogr 12: 361-371. doi: 10.1046/j.1466-822X.2003.00042.x
|
| [21] |
Elith J, Phillips SJ, Hastie T, et al. (2011) A statistical explanation of MaxEnt for ecologists. Divers Distrib 17: 43-57. doi: 10.1111/j.1472-4642.2010.00725.x
|
| [22] | Huntley RE, Green RE, Collinghan Y, et al. (2007) A climatic atlas of European breeding birds. Durham, Sandy and Barcelona: Durham University, 2007. |
| [23] |
Barbet-Massin M, Thuiller W, Jiguet F (2012) The fate of European breeding birds under climate, land-use and dispersal scenarios. Glob Change Biol 18: 881-890. doi: 10.1111/j.1365-2486.2011.02552.x
|
| [24] |
Bucklin DN, Basille M, Benscoter AM, et al. (2015) Comparing species distribution models constructed with different subsets of environmental predictors. Divers Distrib 21: 23-35. doi: 10.1111/ddi.12247
|
| [25] |
Elith J, Leathwick JR (2009) Species Distribution Models: Ecological explanation and prediction across space and time. Annu Rev Ecol Evol Syst 40: 677-697. doi: 10.1146/annurev.ecolsys.110308.120159
|
| [26] | BISON, 2016. Available from: http://bison.usgs.ornl.gov/#home |
| [27] | ORNIS, 2016. Available from: http://www.ornisnet.org/ |
| [28] | VertNet, 2016. Available from: http://portal.vertnet.org/search |
| [29] | GAP Analysis, 2016. Available from: http://gapanalysis.usgs.gov/species/viewer/species-viewer/ |
| [30] | ArcGIS for Desktop, 2016. Available from: http://www.esri.com/software/arcgis/arcgis-for-desktop |
| [31] |
Hijmans RJ, Cameron SE, Parra JL, et al. (2005) Very high resolution interpolated climate surfaces for global land areas. Int J Climatol 25: 1965-1978. doi: 10.1002/joc.1276
|
| [32] |
Dormann CF (2013) Collinearity: a review of methods to deal with it and a simulation study evaluating their performance. Ecography 36: 27-46. doi: 10.1111/j.1600-0587.2012.07348.x
|
| [33] | Gama M, Crespo D, Dolbeth M, et al. (2015) Predicting global habitat suitability for Corbicula fluminea using species distribution models: The importance of different environmental datasets. Ecol Model 319: 163-169. |
| [34] | Talbert C (2012) Software for Assisted Habitat Modeling Package for VisTrails (SAHM: VisTrails). Fort Collins, Colorado, USA: U.S. Geological Survey. |
| [35] |
Poiani KA, Johnson WC (1991) Global warming and prairie wetlands: potential consequences for waterfowl habitat. BioScience 41: 611-618. doi: 10.2307/1311698
|
| [36] |
Araújo MB, New M (2007) Ensemble forecasting of species distributions. Trends Ecol Evol 22: 42-47. doi: 10.1016/j.tree.2006.09.010
|
| [37] |
Breiman L (2001) Random forests. Mach Learn 45: 5-32. doi: 10.1023/A:1010933404324
|
| [38] | Liaw A, Wiener M (2002) Classification and regression by randomForest. R News 2: 18-22. |
| [39] |
Elith J, Leathwick JR, Hastie T (2008) A working guide to boosted regression trees. J Anim Ecol 77: 802-813. doi: 10.1111/j.1365-2656.2008.01390.x
|
| [40] | Phillips SJ, Anderson RP, Schapire RE (2006) Maximum entropy modeling of species geographic distributions. Ecol Model 190: 231-259. |
| [41] |
Phillips SJ, Dudík M (2008) Modeling of species distributions with Maxent: new extensions and a comprehensive evaluation. Ecography 31: 161-175. doi: 10.1111/j.0906-7590.2008.5203.x
|
| [42] |
Leathwick JR, Elith J, Hastie T (2006) Comparative performance of generalized additive models and multivariate adaptive regression splines for statistical modelling of species distributions. Ecol Model 199: 188-196. doi: 10.1016/j.ecolmodel.2006.05.022
|
| [43] |
Elith J (2006) Novel methods improve prediction of species' distributions from occurrence data. Ecography 29: 129-151. doi: 10.1111/j.2006.0906-7590.04596.x
|
| [44] |
Morisette JT (2013) VisTrails SAHM: visualization and workflow management for species habitat modeling. Ecography 36: 129-135. doi: 10.1111/j.1600-0587.2012.07815.x
|
| [45] |
Phillips SJ, Dudík M, Elith J, et al. (2009) Sample selection bias and presence-only distribution models: implications for background and pseudo-absence data. Ecol Appl 19: 181-197. doi: 10.1890/07-2153.1
|
| [46] | Species Viewer. SGS National Gap Analysis Program, 2016. Available from: http://gapanalysis.usgs.gov/species/viewer/species-viewer/ |
| [47] | Willey D, Riper CV (2000) First-year movements by juvenile Mexican Spotted Owls in the Canyonlands of Utah. J Raptor Res: 1-7. |
| [48] |
Campbell H, Harris BK (1965) Mass population dispersal and long-distance movements in Scaled Quail. J Wildl Manag 29: 801-805. doi: 10.2307/3798556
|
| [49] |
VanDerWal J, Shoo LP, Graham C, et al. (2009) Selecting pseudo-absence data for presence-only distribution modeling: How far should you stray from what you know? Ecol Model 220: 589-594. doi: 10.1016/j.ecolmodel.2008.11.010
|
| [50] |
Liu C, Berry PM, Dawson TP, et al. (2005) Selecting thresholds of occurrence in the prediction of species distributions. Ecography 28: 385-393. doi: 10.1111/j.0906-7590.2005.03957.x
|
| [51] | Lobo JM, Jiménez-Valverde A, Real R (2008) AUC: a misleading measure of the performance of predictive distribution models. Glob Ecol Biogeogr 17: 145-151. |
| [52] |
Stohlgren TJ (2010) Ensemble habitat mapping of invasive plant species. Risk Anal 30: 224-235. doi: 10.1111/j.1539-6924.2009.01343.x
|
| [53] | Manel S, Williams HC, Ormerod SJ (2001) Evaluating presence–absence models in ecology: the need to account for prevalence. J Appl Ecol 38: 921-931. |
| [54] |
Rehfeldt GE, Crookston NL, Sáenz-Romero C, et al. (2012) North American vegetation model for land-use planning in a changing climate: a solution to large classification problems. Ecol Appl 22: 119-141. doi: 10.1890/11-0495.1
|
| [55] | Stebbins RC (2003) A Field Guide to Western Reptiles and Amphibians, 3 edition. Boston: Houghton Mifflin Harcourt. |
| [56] |
Warren DL, Seifert SN (2011) Ecological niche modeling in Maxent: the importance of model complexity and the performance of model selection criteria. Ecol Appl 21: 335-342. doi: 10.1890/10-1171.1
|
| [57] |
Allouche O, Tsoar A, Kadmon R (2006) Assessing the accuracy of species distribution models: prevalence, kappa and the true skill statistic (TSS). J Appl Ecol 43: 1223-1232. doi: 10.1111/j.1365-2664.2006.01214.x
|
| [58] | Cohen J (1960) A coefficient of agreement for nominal scales. Educ Psychol Meas 20: 37-46. |
| [59] |
Swets JA (1988) Measuring the accuracy of diagnostic systems. Science 240: 1285-1293. doi: 10.1126/science.3287615
|
| [60] |
Coetzee BWT, Robertson MP, Erasmus BFN, et al. (2009) Ensemble models predict important bird areas in southern Africa will become less effective for conserving endemic birds under climate change. Glob Ecol Biogeogr 18: 701-710. doi: 10.1111/j.1466-8238.2009.00485.x
|
| [61] | Taylor KE, Stouffer RJ, Meehl GA (2011) An Overview of CMIP5 and the experiment design. Bull Am Meteorol Soc 93: 485-498. |
| [62] | Sheffield J (2013) North American Climate in CMIP5 Experiments. Part I: Evaluation of historical simulations of continental and regional climatology. J Clim 26: 9209-9245. |
| [63] |
Gent PR (2011) The Community Climate System Model Version 4. J Clim 24: 4973-4991. doi: 10.1175/2011JCLI4083.1
|
| [64] |
Collins WJ (2011) Development and evaluation of an Earth-System model-HadGEM2. Geosci Model Dev 4: 1051-1075. doi: 10.5194/gmd-4-1051-2011
|
| [65] | Watanabe M. Improved Climate Simulation by MIROC5: Mean States, Variability, and Climate Sensitivity. J Clim 23: 6312-6335. |
| [66] |
Block K, Mauritsen T (2013) Forcing and feedback in the MPI-ESM-LR coupled model under abruptly quadrupled CO2. J Adv Model Earth Syst 5: 676-691. doi: 10.1002/jame.20041
|
| [67] | Roeckner E, Giorgetta MA, Crueger T, et al. (2010) Historical and future anthropogenic emission pathways derived from coupled climate–carbon cycle simulations. Clim Change 105: 91-108. |
| [68] | Arora VK (2011) Carbon emission limits required to satisfy future representative concentration pathways of greenhouse gases. Geophys Res Lett 38: L05805. |
| [69] | Chalmers N, Highwood EJ, Hawkins E, et al. (2012) Aerosol contribution to the rapid warming of near-term climate under RCP 2.6. Geophys Res Lett 39: L18709. |
| [70] |
Vuuren DP (2011) The representative concentration pathways: an overview. Clim Change 109: 5-31. doi: 10.1007/s10584-011-0148-z
|
| [71] | Knutti R, Sedláček J (2013) Robustness and uncertainties in the new CMIP5 climate model projections. Nat Clim Change 3: 369-373. |
| [72] |
Hirzel AH, Le Lay G, Helfer V, et al. (2006) Evaluating the ability of habitat suitability models to predict species presences. Ecol Model 199: 142-152. doi: 10.1016/j.ecolmodel.2006.05.017
|
| [73] |
Kumar S, Spaulding SA, Stohlgren TJ, et al. (2009) Potential habitat distribution for the freshwater diatom Didymosphenia geminata in the continental US. Front Ecol Environ 7: 415-420. doi: 10.1890/080054
|
| [74] |
Capinha C, Anastácio P (2011) Assessing the environmental requirements of invaders using ensembles of distribution models. Divers Distrib 17: 13-24. doi: 10.1111/j.1472-4642.2010.00727.x
|
| [75] |
Robert K, Jones DOB, Roberts JM, et al. (2016) Improving predictive mapping of deep-water habitats: Considering multiple model outputs and ensemble techniques. Deep Sea Res Part Oceanogr Res Pap 113: 80-89. doi: 10.1016/j.dsr.2016.04.008
|
| [76] |
Marmion M, Parviainen M, Luoto M, et al. (2009) Evaluation of consensus methods in predictive species distribution modelling. Divers Distrib 15: 59-69. doi: 10.1111/j.1472-4642.2008.00491.x
|
| [77] |
Poulos HM, Chernoff B, Fuller PL, et al. (2012) Ensemble forecasting of potential habitat for three invasive fishes. Aquat Invasions 7: 59-72. doi: 10.3391/ai.2012.7.1.007
|
| [78] |
Du P, Xia J, Zhang W, et al. (2012) Multiple classifier system for remote sensing image classification: A review. Sensors 12: 4764-4792. doi: 10.3390/s120404764
|
| [79] |
Meinshausen M (2011) The RCP greenhouse gas concentrations and their extensions from 1765 to 2300. Clim Change 109: 213. doi: 10.1007/s10584-011-0156-z
|
| [80] | Barsugli JJ (2013) The practitioner's dilemma: How to assess the credibility of downscaled climate projections. Eos Trans Am Geophys Union 94: 424-425. |
| [81] |
Dixon KW (2016) Evaluating the stationarity assumption in statistically downscaled climate projections: is past performance an indicator of future results? Clim Change 135: 395-408. doi: 10.1007/s10584-016-1598-0
|
| [82] |
Stoklosa J, Daly C, Foster SD, et al. (2015) A climate of uncertainty: accounting for error in climate variables for species distribution models. Methods Ecol Evol 6: 412-423. doi: 10.1111/2041-210X.12217
|
| [83] |
Bakkenes M, Alkemade JRM, Ihle F, et al. (2002) Assessing effects of forecasted climate change on the diversity and distribution of European higher plants for 2050. Glob Change Biol 8: 390-407. doi: 10.1046/j.1354-1013.2001.00467.x
|
| [84] |
Bateman BL, Murphy HT, Reside AE, et al. (2013) Appropriateness of full-, partial- and no-dispersal scenarios in climate change impact modelling. Divers Distrib 19: 1224-1234. doi: 10.1111/ddi.12107
|
| [85] | U.S. Fish and Wildlife Service, Mexican spotted owl (Strix occidentalis lucida) 5-Year review short form summary. Arizona Ecological Services Office, Arizona, USA, 2013. |
| [86] |
Davis DM (2009) Nesting ecology and reproductive success of Lesser Prairie-Chickens in shinnery oak-dominated rangelands. Wilson J Ornithol 121: 322-327. doi: 10.1676/08-090.1
|
| [87] | Jarnevich CS (2016) Assessing range-wide habitat suitability for the Lesser Prairie-Chicken. Avian Conserv Ecol 11: 2. |
| [88] | Jennison R, Pitman J, Kramer J, et al., Prairie Chicken LEK Survey, Kansas Department of Wildlife, Parks, & Tourism, Kansas, USA, 2013. |
| [89] | Greenwald N, Petition to list the white-tailed ptarmigan (Lagopus leucura) as a threatened species under the endangered species act. Center for Biological Diversity, Petition, 2010. |
| [90] | NM Crucial Habitat Assessment Tool, 2016. Available from: http://nmchat.org/. |
| [91] | Species Reports, 2016. Available from: http://ecos.fws.gov/tess_public/. |
| [92] | Endangered Species Program, 2016. Available from: http://www.fws.gov/endangered/species/. |
| [93] | Christmas Bird Count. Audubon, 2016. Available from: https://www.audubon.org/conservation/science/christmas-bird-count. |
| [94] |
Eaton JG, Scheller RM (1996) Effects of climate warming on fish thermal habitat in streams of the United States. Limnol Oceanogr 41: 1109-1115. doi: 10.4319/lo.1996.41.5.1109
|
| [95] | Johnson K, Tonne P, Muldavin E (2004) New Mexico drought at risk species as determined by Natural Heritage New Mexico (NHNM) staff at the University of New Mexico. Museum of Southwestern Biology, Albuquerque, NM, USA, 2004. |
| [96] | Enquist C, Gori D (2008) Implications of recent climate change on conservation priorities in New Mexico. The Nature Conservancy, Santa Fe, NM, USA, 2008. |
| [97] | Mitchell NJ, Janzen FJ (2010) Temperature-dependent sex determination and contemporary climate change. Sex Dev Genet Mol Biol Evol Endocrinol Embryol Pathol Sex Determ Differ 4: 129-140. |
| [98] | La Sorte FA, Jetz W (2010) Projected range contractions of montane biodiversity under global warming. Proc R Soc Lond B Biol Sci: rspb20100612. |
| [99] | Zack S, Ellison K, Cross M, et al., Climate change planning for the Great Plains: Wildlife vulnerability assessment & recommendations for land and grazing management. Wildlife Conservation Society, North America Program, Bozeman, MT, USA, 2010. |
| [100] | Coe SJ, Finch DM, Friggens MM (2012) An assessment of climate change and the vulnerability of wildlife in the Sky Islands of the Southwest. Gen. Tech. Rep. RMRS-GTR-273. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. 208 p. |
| [101] | Bagne KE, Finch DM (2013) Vulnerability of species to climate change in the Southwest: threatened, endangered, and at-risk species at Fort Huachuca, Arizona. |
| [102] | Friggens MM, Finch DM, Bagne KE, et al. (2013) Vulnerability of species to climate change in the Southwest: terrestrial species of the Middle Rio Grande. Gen. Tech. Rep. RMRS-GTR-306. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. 191 p. |
| [103] |
Moyle PB, Kiernan JD, Crain PK, et al. (2013) Climate Change Vulnerability of Native and Alien Freshwater Fishes of California: A Systematic Assessment Approach. PLOS ONE 8: e63883. doi: 10.1371/journal.pone.0063883
|
| [104] | Species Data and Modeling, 2016. Available from: http://gapanalysis.usgs.gov/species/data/. |
| [105] | IUCN Red List of Threatened Species, 2016. Available from: http://www.iucnredlist.org/. |
| [106] | Nature Serve Explorer, 2016. Available from: http://explorer.natureserve.org/. |
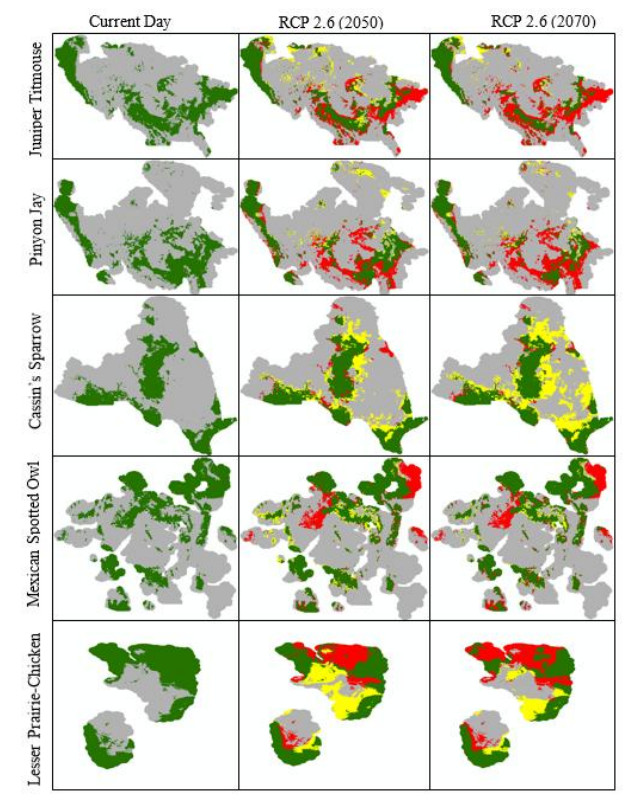
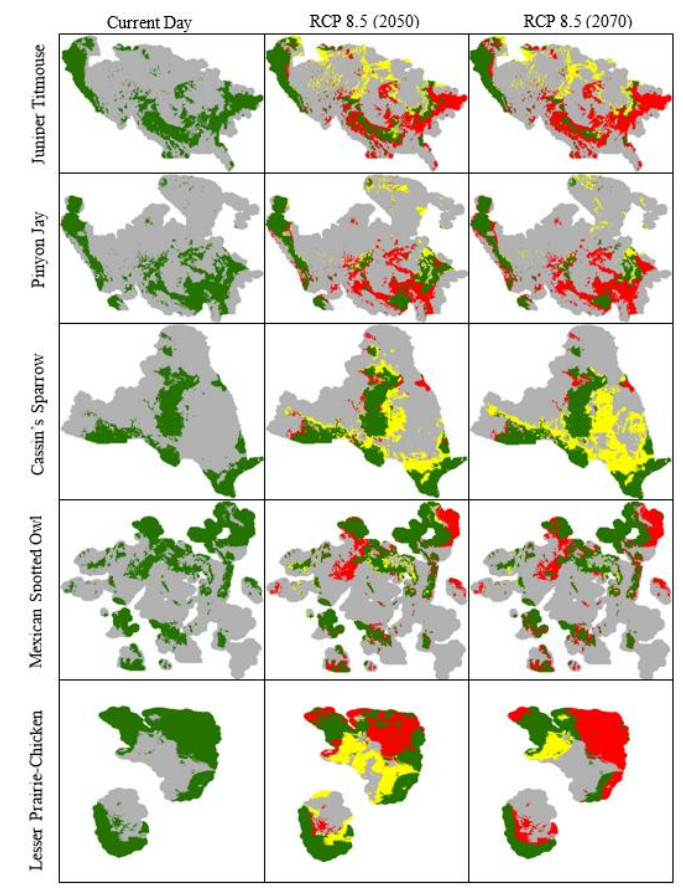
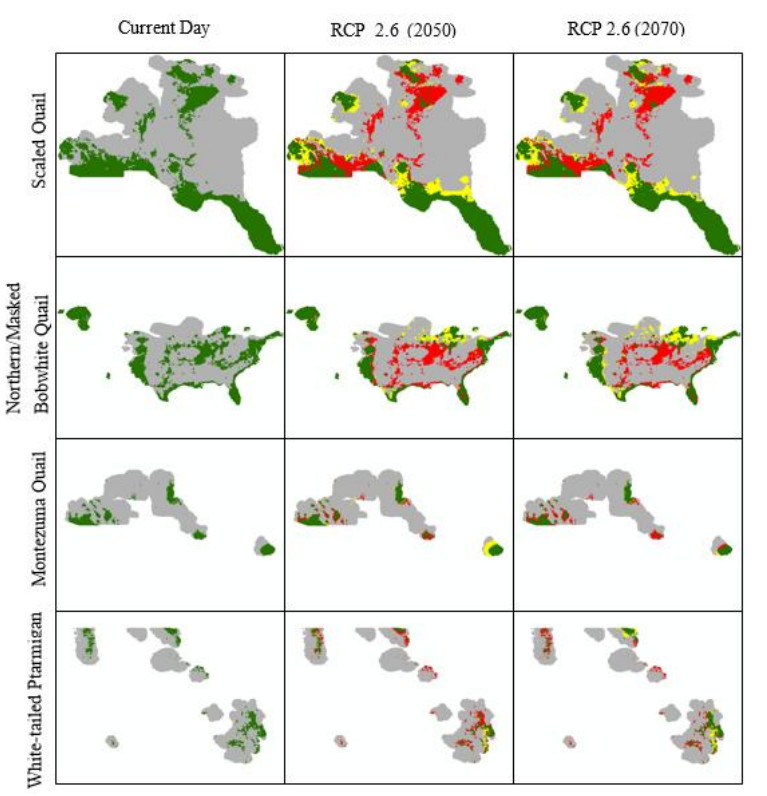


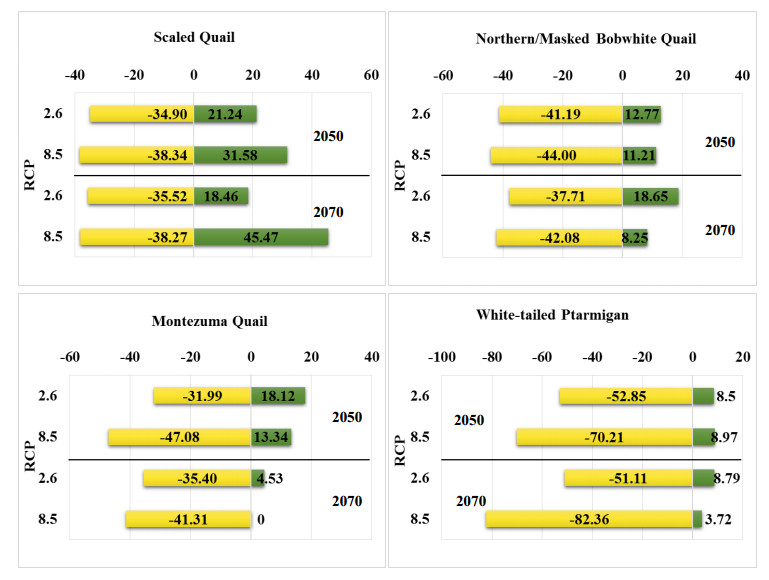
Figures(10) / Tables(4)

Eric Ariel L. Salas, Virginia A. Seamster, Kenneth G. Boykin, Nicole M. Harings, Keith W. Dixon. Modeling the impacts of climate change on Species of Concern (birds) in South Central U.S. based on bioclimatic variables[J]. AIMS Environmental Science, 2017, 4(2): 358-385. doi: 10.3934/environsci.2017.2.358







 DownLoad:
DownLoad: