




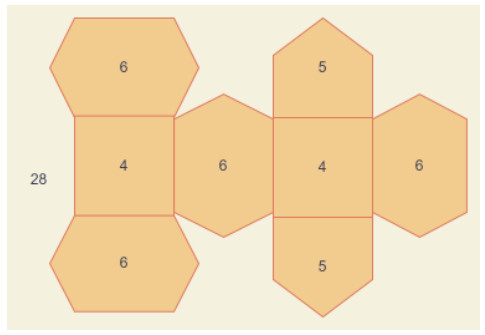
Figure 1.
An example of face degree.
Citation: Gérald Gaibelet, François Tercé, Sophie Allart, Chantal Lebrun, Xavier Collet, Nadège Jamin, Stéphane Orlowski. Fluorescent probes for detecting cholesterol-rich ordered membrane microdomains: entangled relationships between structural analogies in the membrane and functional homologies in the cell[J]. AIMS Biophysics, 2017, 4(1): 121-151. doi: 10.3934/biophy.2017.1.121
| [1] | Huilin Ge, Yuewei Dai, Zhiyu Zhu, Biao Wang . Robust face recognition based on multi-task convolutional neural network. Mathematical Biosciences and Engineering, 2021, 18(5): 6638-6651. doi: 10.3934/mbe.2021329 |
| [2] | Chii-Dong Ho, Gwo-Geng Lin, Thiam Leng Chew, Li-Pang Lin . Conjugated heat transfer of power-law fluids in double-pass concentric circular heat exchangers with sinusoidal wall fluxes. Mathematical Biosciences and Engineering, 2021, 18(5): 5592-5613. doi: 10.3934/mbe.2021282 |
| [3] | José M. Sigarreta . Extremal problems on exponential vertex-degree-based topological indices. Mathematical Biosciences and Engineering, 2022, 19(7): 6985-6995. doi: 10.3934/mbe.2022329 |
| [4] | Qingqun Huang, Muhammad Labba, Muhammad Azeem, Muhammad Kamran Jamil, Ricai Luo . Tetrahedral sheets of clay minerals and their edge valency-based entropy measures. Mathematical Biosciences and Engineering, 2023, 20(5): 8068-8084. doi: 10.3934/mbe.2023350 |
| [5] | Hao Wang, Guangmin Sun, Kun Zheng, Hui Li, Jie Liu, Yu Bai . Privacy protection generalization with adversarial fusion. Mathematical Biosciences and Engineering, 2022, 19(7): 7314-7336. doi: 10.3934/mbe.2022345 |
| [6] | Meili Tang, Qian Pan, Yurong Qian, Yuan Tian, Najla Al-Nabhan, Xin Wang . Parallel label propagation algorithm based on weight and random walk. Mathematical Biosciences and Engineering, 2021, 18(2): 1609-1628. doi: 10.3934/mbe.2021083 |
| [7] | Xinmei Liu, Xinfeng Liang, Xianya Geng . Expected Value of Multiplicative Degree-Kirchhoff Index in Random Polygonal Chains. Mathematical Biosciences and Engineering, 2023, 20(1): 707-719. doi: 10.3934/mbe.2023032 |
| [8] | Fengwei Li, Qingfang Ye, Juan Rada . Extremal values of VDB topological indices over F-benzenoids with equal number of edges. Mathematical Biosciences and Engineering, 2023, 20(3): 5169-5193. doi: 10.3934/mbe.2023240 |
| [9] | Stefano Cosenza, Paolo Crucitti, Luigi Fortuna, Mattia Frasca, Manuela La Rosa, Cecilia Stagni, Lisa Usai . From Net Topology to Synchronization in HR Neuron Grids. Mathematical Biosciences and Engineering, 2005, 2(1): 53-77. doi: 10.3934/mbe.2005.2.53 |
| [10] | Qiming Li, Tongyue Tu . Large-pose facial makeup transfer based on generative adversarial network combined face alignment and face parsing. Mathematical Biosciences and Engineering, 2023, 20(1): 737-757. doi: 10.3934/mbe.2023034 |
To exemplify the phenomena of compounds scientifically, researchers utilize the contraption of the diagrammatic hypothesis, it is a well-known branch of geometrical science named graph theory. This division of numerical science provides its services in different fields of sciences. The particular example in networking [1], from electronics [2], and for the polymer industry, we refer to see [3]. Particularly in chemical graph theory, this division has extra ordinary assistance to study giant and microscope-able chemical compounds. For such a study, researchers made some transformation rules to transfer a chemical compound to a discrete pattern of shapes (graph). Like, an atom represents as a vertex and the covalent bonding between atoms symbolized as edges. Such transformation is known as molecular graph theory. A major importance of this alteration is that the hydrogen atoms are omitted. Some chemical structures and compounds conversion are presented in [4,5,6].
In cheminformatics, the topological index gains attraction due to its implementations. Various topological indices help to estimate a bio-activity and physicochemical characteristics of a chemical compound. Some interesting and useful topological indices for various chemical compounds are studied in [3,7,8]. A topological index modeled a molecular graph or a chemical compound into a numerical value. Since 1947, topological index implemented in chemistry [9], biology [10], and information science [11,12]. Sombor index and degree-related properties of simplicial networks [13], Nordhaus–Gaddum-type results for the Steiner Gutman index of graphs [14], Lower bounds for Gaussian Estrada index of graphs [15], On the sum and spread of reciprocal distance Laplacian eigenvalues of graphs in terms of Harary index [16], the expected values for the Gutman index, Schultz index, and some Sombor indices of a random cyclooctane chain [17,18,19], bounds on the partition dimension of convex polytopes [20,21], computing and analyzing the normalized Laplacian spectrum and spanning tree of the strong prism of the dicyclobutadieno derivative of linear phenylenes [22], on the generalized adjacency, Laplacian and signless Laplacian spectra of the weighted edge corona networks [23,24], Zagreb indices and multiplicative Zagreb indices of Eulerian graphs [25], Minimizing Kirchhoff index among graphs with a given vertex bipartiteness, [26], asymptotic Laplacian energy like invariant of lattices [27]. Few interesting studies regarding the chemical graph theory can be found in [28,29,30,31,32].
Recently, the researchers of [33] introduced a topological descriptor and called the face index. Moreover, the idea of computing structure-boiling point and energy of a structure, motivated them to introduced this parameter without heavy computation. They computed these parameters for different models compare the results with previous literature and found approximate solutions with comparatively less computations. This is all the blessings of face index of a graph. The major concepts of this research work are elaborated in the given below definitions.
Definition 1.1. [33] Let a graph G=(V(G),E(G),F(G)) having face, edge and vertex sets notation with F(G),E(G),V(G), respectively. It is mandatory that the graph is connected, simple and planar. If e from the edge set E(G), is one of those edges which surrounds a face, then the face f from the face set F(G), is incident to the edge e. Likewise, if a vertex α from the vertex set V(G) is at the end of those incident edges, then a face f is incident to that vertex. This face-vertex incident relation is symbolized here by the notation α∼f. The face degree of f in G is described as d(f)=∑α∼fd(α), which are elaborated in the Figure 1.
Definition 1.2. [33] The face index FI(G), for a graph G, is formulated as
| FI(G)=∑f∈F(G)d(f)=∑α∼f,f∈F(G)d(α). |
In the Figure 1, we can see that there are two faces with degree 4, exactly two with five count and four with count of 6. Moreover, there is an external face with count of face degree 28, which is the count of vertices.
As the information given above that the face index is quite new and introduced in the year 2020, so there is not so much literature is available. A few recent studies on this topic are summarized here. A chemical compound of silicon carbides is elaborated with such novel definition in [34]. Some carbon nanotubes are discussed in [35]. Except for the face index, there are distance and degree-based graphical descriptors available in the literature. For example, distance-based descriptors of phenylene nanotube are studied in [36], and in [37] titania nanotubes are discussed with the same concept. Star networks are studied in [38], with the concept of degree-based descriptors. Bounds on the descriptors of some generalized graphs are discussed in [39]. General Sierpinski graph is discussed in [40], in terms of different topological descriptor aspects. The study of hyaluronic acid-doxorubicin ar found in [41], with the same concept of the index. The curvilinear regression model of the topological index for the COVID-19 treatment is discussed in [42]. For further reading and interesting advancements of topological indices, polynomials of zero-divisor structures are found in [43], zero divisor graph of commutative rings [44], swapped networks modeled by optical transpose interconnection system [45], metal trihalides network [46], some novel drugs used in the cancer treatment [47], para-line graph of Remdesivir used in the prevention of corona virus [48], tightest nonadjacently configured stable pentagonal structure of carbon nanocones [49]. In order to address a novel preventive category (P) in the HIV system known as the HIPV mathematical model, the goal of this study is to offer a design of a Morlet wavelet neural network (MWNN) [50].
In the next section, we discussed the newly developed face index or face-based index for different chemical compounds. Silicate network, triangular honeycomb network, carbon sheet, polyhedron generalized sheet, and generalized chain of silicate network are studied with the concept of the face-based index. Given that the face index is more versatile than vertex degree-based topological descriptors, this study will aid in understanding the structural characteristics of chemical networks. Only the difficulty authors will face to compute the face degree of a generalized network or structure, because it is more generalized version and taking degree based partition of edges into this umbrella of face index.
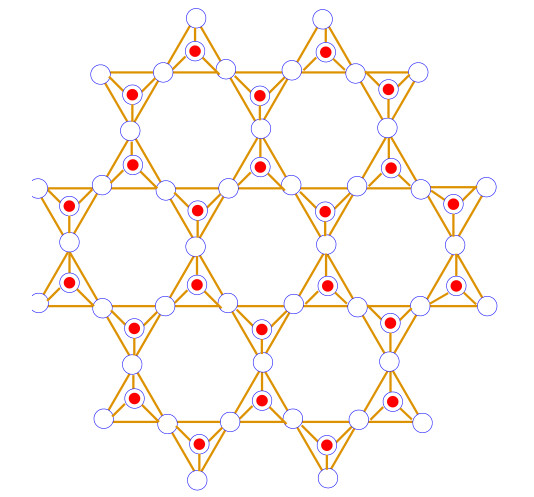
Silicates are formed when metal carbonates or metal oxides react with sand. The SiO4, which has a tetrahedron structure, is the fundamental chemical unit of silicates. The central vertex of the SiO4 tetrahedron is occupied by silicon ions, while the end vertices are occupied by oxygen ions [51,52,53]. A silicate sheet is made up of rings of tetrahedrons that are joined together in a two-dimensional plane by oxygen ions from one ring to the other to form a sheet-like structure. The silicate network SLn symbol, where n represents the total number of hexagons occurring between the borderline and center of the silicate network SLn. The silicate network of dimension one is depicted in Figure 2. It contain total 3n(5n+1) vertices are 36n2 edges. Moreover, the result required is detailed are available in Table 1.
| Dimension | |f12| | |f15| | |f36| |
| 1 | 24 | 48 | 7 |
| 2 | 32 | 94 | 14 |
| 3 | 40 | 152 | 23 |
| 4 | 48 | 222 | 34 |
| 5 | 56 | 304 | 47 |
| 6 | 64 | 398 | 62 |
| 7 | 72 | 504 | 79 |
| 8 | 80 | 622 | 98 |
| . | . | . | . |
| . | . | . | . |
| . | . | . | . |
| n | 8n+16 | 6n2+28n+14 | n2+4n+2 |
 DownLoad:
CSV
DownLoad:
CSV
Theorem 2.1. Let SLn be the silicate network of dimension n≥1. Then the face index of SLn is
| FI(SLn)=126n2+720n+558. |
Proof. Consider SLn the graph of silicate network with dimension n. Suppose fi denotes the faces of graph SLn having degree i. that is, d(fi)=∑α∼fid(α)=i and |fi| denotes the number of faces with degree i. The graph SLn contains three types of internal faces f12, f15, f36, and single external face which is usually denoted by f∞.
If SLn has one dimension then sum of degree of vertices incident to the external face is 144 and when SLn has two dimension then sum of degree of incident vertices to the external face is 204 whenever SLn has three dimension then sum of degree of incident vertices to the external face is 264. Similarly, SLn has n−dimension then sum of degree of incident vertices to the external face is 60n+84.
The number of internal faces with degree in each dimension is mentioned in Table 1.
By using the definition of face index FI we have
| FI(SLn)=∑α∼f∈F(SLn)d(α)=∑α∼f12∈F(SLn)d(α)+∑α∼f15∈F(SLn)d(α)+∑α∼f36∈F(SLn)d(α)+∑α∼f∞∈F(SLn)d(α)=|f12|(12)+|f15|(15)+|f36|(36)+(60n+84)=(8n+16)(12)+(6n2+28n+14)(15)+(n2+4n+2)(36)+60n+84=126n2+72n+558. |
Hence, this is our required result.
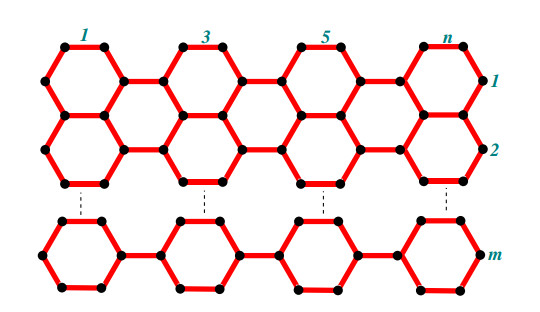
A chain silicate network of dimension (m,n) is symbolized as CSL(m,n) which is made by arranging (m,n) tetrahedron molecules linearly. A chain silicate network of dimension (m,n) with m,n≥1 where m denotes the number of rows and each row has n number of tetrahedrons. The following theorem formulates the face index FI for chain silicate network.
Theorem 2.2. Let CSL(m,n) be the chain of silicate network of dimension m,n≥1. Then the face index FI of the graph CSL(m,n) is
| FI(CSL(m,n))={48n−12if m=1, n≥1;96m−12if n=1, m≥2;168m−60if n=2,m≥2;45m−9n+36mn−42if both m,n are even45m−9n+36mn−21otherwise. |
Proof. Let CSL(m,n) be the graph of chain silicate network of dimension (m,n) with m,n≥1 where m represents the number of rows and n is the number of tetrahedrons in each row. A graph CSL(m,n) for m=1 contains three type of internal faces f9, f12 and f15 with one external face f∞. While for m≥2, it has four type of internal faces f9, f12, f15 and f36 with one external face f∞. We want to evaluate the algorithm of face index FI for chain silicate network. We will discuss it in two different cases.
Case 1: When CSL(m,n) has one row (m=1) with n number of tetrahedrons as shown in the Figure 3.
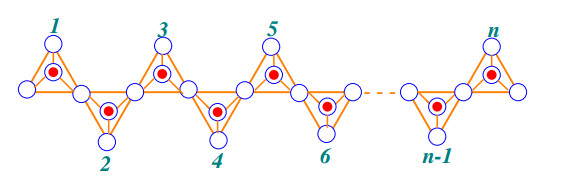
The graph has three type of internal faces f9, f12 and f15 with one external face f∞. The sum of degree of incident vertices to the external face is 9n and number of faces are |f9|=2, |f12|=2n and |f15|=n−2. Now the face index FI of the graph CSL(m,n) is given by
| FI(CSL(m,n))=∑α∼f∈F(CSL(m,n))d(α)=∑α∼f9∈F(CSL(m,n))d(α)+∑α∼f12∈F(CSL(m,n))d(α)+∑α∼f15∈F(CSL(m,n))d(α)+∑α∼f∞∈F(CSL(m,n))d(α)=|f9|(9)+|f12|(12)+|f15|(15)+(9n)=(2)(9)+(2n)(12)+(n−2)(15)+9n=48n−12. |
Case 2: When CSL(m,n) has more than one rows (m≠1) with n number of tetrahedrons in each row as shown in the Figure 4.
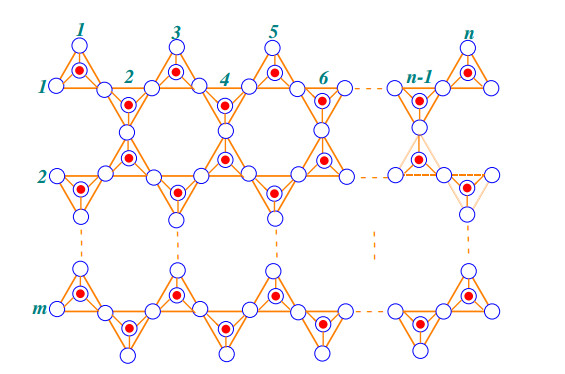
The graph has four type of internal faces f9, f12, f15 and f36 with one external face f∞. The sum of degree of incident vertices to the external face is
| ∑α∼f∞∈F(CSL(m,n))d(α)={18mif n=1, m≥1;27mif n=2, m≥1;30m+15n−30if both m,n are even30m+15n−33otherwise. |
The number of faces are |f9|, |f12|, f15 and |f36| are given by
| |f9|={2if m is odd3+(−1)nif m is even.|f12|={2(2m+n−1)if m is odd4(⌊n+12⌋+2m−1)if m is even|f15|=(3m−2)n−m|f36|={(m−12)(n−1)if m is odd(2n+(−1)n−14)(m−22)nif m is even. |
Now the face index FI of the graph CSL(m,n) is given by
| FI(CSL(m,n))=∑α∼f∈F(CSL(m,n))d(α)=∑α∼f9∈F(CSL(m,n))d(α)+∑α∼f12∈F(CSL(m,n))d(α)+∑α∼f15∈F(CSL(m,n))d(α)+∑α∼f36∈F(CSL(m,n))d(α)+∑α∼f∞∈F(CSL(m,n))d(α)=|f9|(9)+|f12|(12)+|f15|(15)+|f36|(36)+∑α∼f∞∈F(CSL(m,n))d(α). |
After some mathematical simplifications, we can get
| FI(CSL(m,n))={48n−12if m=196m−12if n=1,∀m168m−60if n=2,∀m45m−9n+36mn−42if both m,n are even45m−9n+36mn−21otherwise. |
There are three regular plane tessellations known to exist, each constituted from the same type of regular polygon: triangular, square, and hexagonal. The triangular tessellation is used to define the hexagonal network, which is extensively studied in [54]. A dimensioned hexagonal network THk has 3k2−3k+1 vertices and 9k2−15k+6 edges, where k is the number of vertices on one side of the hexagon. It has 2k−2 diameter. There are six vertices of degree three that are referred to as corner vertices. Moreover, the result required detailed are available in the Table 2.
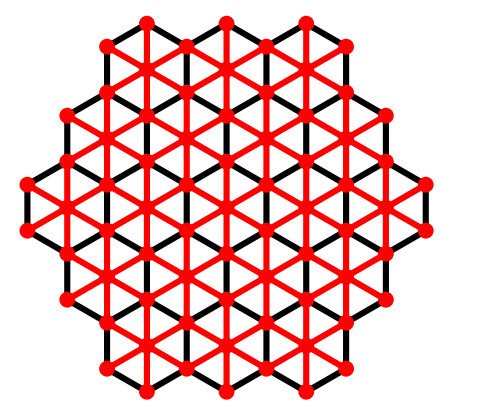
| Dimension | |f12| | |f14| | |f17| | |f18| |
| 1 | 6 | 0 | 0 | 0 |
| 2 | 6 | 12 | 12 | 12 |
| 3 | 6 | 24 | 24 | 60 |
| 4 | 6 | 36 | 36 | 144 |
| 5 | 6 | 48 | 48 | 264 |
| 6 | 6 | 60 | 60 | 420 |
| 7 | 6 | 72 | 72 | 612 |
| 8 | 6 | 84 | 84 | 840 |
| . | . | . | . | . |
| . | . | . | . | . |
| . | . | . | . | . |
| k | 6 | 12(k−1) | 12(k−1) | 18k2−42k+24 |
DownLoad:
CSV
Theorem 2.3. Let THk be the triangular honeycomb network of dimension k≥1. Then the face index of graph THk is
| FI(THk)=324k2−336k+102. |
Proof. Consider THk be a graph of triangular honeycomb network. The graph TH1 has one internal and only one external face while graph THk with k≥2, contains four types of internal faces f12, f14, f17, and f18 with one external face f∞.
For TH1 the sum of degree of incident vertices to the external face is 18 and in TH2 the sum of degree of incident vertices to the external face is 66. Whenever the graph TH3, the sum of degree of incident vertices to the external face is 114. Similarly, for THk has n−dimension then sum of degree of incident vertices to the external face is 48k−30.
The number of internal faces with degree in each dimension is given in Table 2.
By using the definition of face index FI we have
| FI(THk)=∑α∼f∈F(THk)d(α)=∑α∼f12∈F(THk)d(α)+∑α∼f14∈F(THk)d(α)+∑α∼f17∈F(THk)d(α)+∑α∼f18∈F(THk)d(α)+∑α∼f∞∈F(THk)d(α)=|f12|(12)+|f14|(14)+|f17|(17)+|f18|(18)+(48k−30)=(6)(12)+(12(k−1))(14)+(12(k−1))(17)+(18k2−42k+24)(18)+48k−30=324k2−336k+102. |
Hence, this is our required result.
Given carbon sheet in the Figure 6, is made by grid of hexagons. There are few types of carbon sheets are given in [55,56]. The carbon sheet is symbolize as HCSm,n, where n represents the total number of vertical hexagons and m denotes the horizontal hexagons. It contain total 4mn+2(n+m)−1 vertices and 6nm+2m+n−2 edges. Moreover, the result required detailed are available in Tables 3 and 4.
| Dimension m | |f15| | |f16| | |f18| | |f∞| |
| 2 | 3 | 2(n−1) | n−1 | 20n+7 |
DownLoad:
CSV
| Dimension m | |f15| | |f16| | |f17| | |f18| | |f∞| |
| 2 | 3 | 2(n−1) | 0 | n−1 | 20n+7 |
| 3 | 2 | 2n | 1 | 3(n−1) | 20n+17 |
| 4 | 2 | 2n | 3 | 5(n−1) | 20n+27 |
| 5 | 2 | 2n | 5 | 7(n−1) | 20n+37 |
| 6 | 2 | 2n | 7 | 9(n−1) | 20n+47 |
| . | . | . | . | . | . |
| . | . | . | . | . | . |
| . | . | . | . | . | . |
| m | 2 | 2n | 2m−5 | 2mn−2m−3n+3 | 20n+10m−13 |
DownLoad:
CSV
Theorem 2.4. Let HCSm,n be the carbon sheet of dimension (m,n) and m,n≥2. Then the face index of HCSm,n is
| FI(HCSm,n)={70n+2ifm=236mn−14−2(n−4m)ifm≥3. |
Proof. Consider HCSm,n be the carbon sheet of dimension (m,n) and m,n≥2. Let fi denotes the faces of graph HCSm,n having degree i, which is d(fi)=∑α∼fid(α)=i, and |fi| denotes the number of faces with degree i. A graph HCSm,n for a particular value of m=2 contains three types of internal faces f15, f16, f17 and f18 with one external face f∞. While for the generalize values of m≥3, it contain four types of internal faces f15, f16 and f17 with one external face f∞ in usual manner. For the face index of generalize nanotube, we will divide into two cases on the values of m.
Case 1: When HCSm,n has one row or HCS2,n.
A graph HCSm,n for a this particular value of m=2 contains three types of internal faces |f15|=3, |f16|=2(n−1) and |f18|=n−1 with one external face f∞. For the face index of carbon sheet, details are given in the Table 3. Now the face index FI of the graph NT2,n is given by
| FI(HCS2,n)=∑α∼f∈F(HCS2,n)d(α)=∑α∼f15∈F(HCS2,n)d(α)+∑α∼f16∈F(HCS2,n)d(α)+∑α∼f18∈F(HCS2,n)d(α)+∑α∼f∞∈F(HCS2,n)d(α)=|f15|(15)+|f16|(16)+|f18|(18)+20n+7.=3(15)+2(n−1)(16)+(n−1)(18)+20n+7.=70n+2. |
Case 2: When HCSm,n has m≥3 rows.
A graph HCSm,n for generalize values of m≥3 contains four types of internal faces |f15|=2, |f16|=2n, |f17|=2m−5 and |f18|=2mn−2m−3n+3 with one external face f∞. For the face index of carbon sheet, details are given in the Table 4. Now the face index FI of the graph NTm,n is given by
| FI(HCSm,n)=∑α∼f∈F(HCSm,n)d(α)=∑α∼f15∈F(HCSm,n)d(α)+∑α∼f16∈F(HCSm,n)d(α)+∑α∼f17∈F(HCSm,n)d(α)+∑α∼f18∈F(HCSm,n)d(α)+∑α∼f∞∈F(HCSm,n)d(α)=|f15|(15)+|f16|(16)+|f17|(17)+|f18|(18)+20n+10m−13.=36mn−2n+8m−14. |
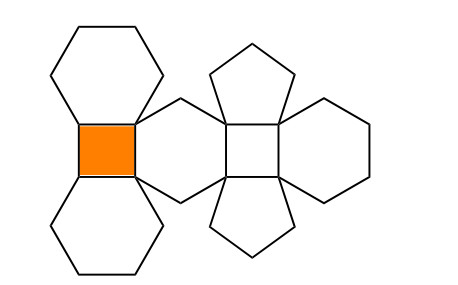
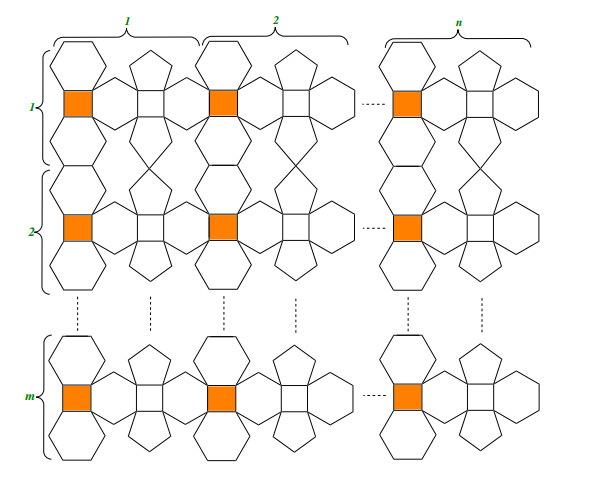
Given structure of polyhedron generalized sheet of C∗28 in the Figure 7, is made by generalizing a C∗28 polyhedron structure which is shown in the Figure 8. This particular structure of C∗28 polyhedron are given in [57]. The polyhedron generalized sheet of C∗28 is as symbolize PHSm,n, where n represents the total number of vertical C∗28 polyhedrons and m denotes the horizontal C∗28 polyhedrons. It contain total 23nm+3n+2m vertices and 33nm+n+m edges. Moreover, the result required detailed are available in Tables 3 and 5.
| m | |f14| | |f15| | |f16| | |f17| | |f18| | |f20| | |f35| |
| 1 | 2n+1 | 2 | 4n−2 | 0 | 0 | 2n−1 | 0 |
| 2 | 2n+2 | 2 | 8n−2 | 2 | 2n−2 | 4n−2 | 2n−1 |
| 3 | 2n+3 | 2 | 12n−2 | 4 | 4n−4 | 6n−3 | 4n−2 |
| . | . | . | . | . | . | . | . |
| . | . | . | . | . | . | . | . |
| . | . | . | . | . | . | . | . |
| m | 2n+m | 2 | 4mn−2 | 2m−2 | 2mn−2(m+n)+2 | 2mn−m | 2mn−(m+2n)+1 |
DownLoad:
CSV
Theorem 2.5. Let PHSm,n be the polyhedron generalized sheet of C∗28 of dimension (m,n) and m,n≥1. Then the face index of PHSm,n is
| FI(PHSm,n)=210mn−2(3m+5n). |
Proof. Consider PHSm,n be the polyhedron generalized sheet of C∗28 of dimension (m,n) and m,n≥1. Let fi denotes the faces of graph PHSm,n having degree i, which is d(fi)=∑α∼fid(α)=i, and |fi| denotes the number of faces with degree i. A graph PHSm,n for the generalize values of m,n≥1, it contain seven types of internal faces f14,f15,f16,f17,f18,f20 and f35 with one external face f∞ in usual manner. For the face index of polyhedron generalized sheet, details are given in the Table 5.
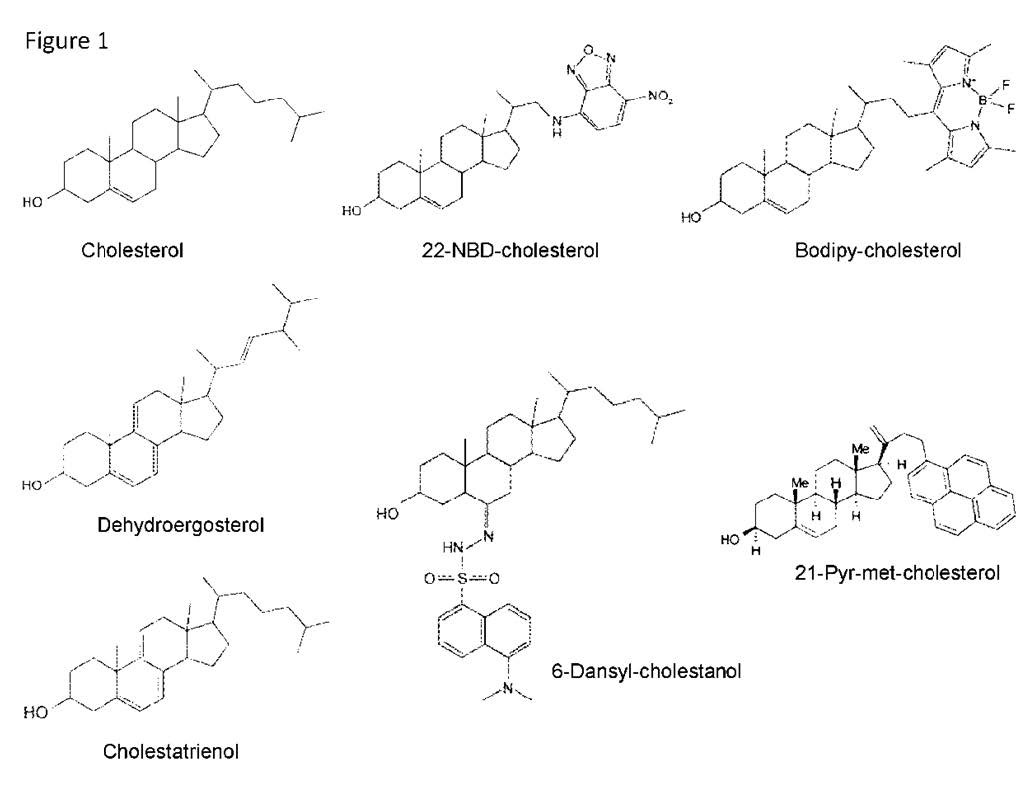
A graph PHSm,n for generalize values of m,n≥1 contains seven types of internal faces |f14|=2n+m, |f15|=2, |f16|=4nm−2, |f17|=2(m−1), |f18|=2nm−2(m+n)+2, |f20|=2nm−2mn−m, and |f35|=2mn−m−2n+1 with one external face f∞. Now the face index FI of the graph PHSm,n is given by
| FI(PHSm,n)=∑α∼f∈F(PHSm,n)d(α)=∑α∼f14∈F(PHSm,n)d(α)+∑α∼f15∈F(PHSm,n)d(α)+∑α∼f16∈F(PHSm,n)d(α)+∑α∼f17∈F(PHSm,n)d(α)+∑α∼f18∈F(PHSm,n)d(α)+∑α∼f20∈F(PHSm,n)d(α)+∑α∼f35∈F(PHSm,n)d(α)+∑α∼f∞∈F(PHSm,n)d(α)=|f14|(14)+|f15|(15)+|f16|(16)+|f17|(17)+|f18|(18)+|f20|(20)+|f35|(35)+37m+68n−35.=210mn−6m−10n. |
With the advancement of technology, types of equipment and apparatuses of studying different chemical compounds are evolved. But topological descriptors or indices are still preferable and useful tools to develop numerical science of compounds. Therefore, from time to time new topological indices are introduced to study different chemical compounds deeply. In this study, we discussed a newly developed tool of some silicate type networks and generalized sheets, carbon sheet, polyhedron generalized sheet, with the face index concept. It provides numerical values of these networks based on the information of faces. It also helps to study physicochemical characteristics based on the faces of silicate networks.
M. K. Jamil conceived of the presented idea. K. Dawood developed the theory and performed the computations. M. Azeem verified the analytical methods, R. Luo investigated and supervised the findings of this work. All authors discussed the results and contributed to the final manuscript.
This work was supported by the National Science Foundation of China (11961021 and 11561019), Guangxi Natural Science Foundation (2020GXNSFAA159084), and Hechi University Research Fund for Advanced Talents (2019GCC005).
The authors declare that they have no conflicts of interest.
| [1] |
Klymchenko AS, Kreder R (2014) Fluorescent probes for lipid rafts: from model membranes to living cells. Chem Biol 21: 97-113. doi: 10.1016/j.chembiol.2013.11.009

|
| [2] |
Sankaram MB, Thompson TE (1991) Cholesterol-induced fluid-phase immiscibility in membranes. Proc Natl Acad Sci USA 88: 8686-8690. doi: 10.1073/pnas.88.19.8686
|
| [3] |
Veatch SL, Keller SL (2005) Seeing spots: complex phase behavior in simple membranes. Biochim Biophys Acta 1746: 172-185. doi: 10.1016/j.bbamcr.2005.06.010
|
| [4] |
Daly TA, Wang M, Regen SL (2011) The origin of cholesterol's condensing effect. Langmuir 27: 2159-2161. doi: 10.1021/la105039q
|
| [5] |
Hakobyan D, Heuer A (2014) Key molecular requirements for raft formation in lipid/cholesterol membranes. PLoS One 9: e87369. doi: 10.1371/journal.pone.0087369
|
| [6] |
Krause MR, Regen SL (2014) The structural role of cholesterol in cell membranes: from condensed bilayers to lipid rafts. Acc Chem Res 47: 3512-3521. doi: 10.1021/ar500260t
|
| [7] |
Feigenson GW (2006) Phase behavior of lipid mixtures. Nat Chem Biol 2: 560-563. doi: 10.1038/nchembio1106-560
|
| [8] |
Rog T, Pasenkiewicz-Gierula M, Vattulainen I, et al. (2009) Ordering effects of cholesterol and its analogues. Biochim Biophys Acta 1788: 97-121. doi: 10.1016/j.bbamem.2008.08.022
|
| [9] |
Garcia-Saez AJ, Schwille P (2010) Stability of lipid domains. FEBS Lett 584: 1653-1658. doi: 10.1016/j.febslet.2009.12.036
|
| [10] |
Shaw JE, Epand RF, Epand RM, et al. (2006) Correlated fluorescence-atomic force microscopy of membrane domains: structure of fluorescence probes determines lipid localization. Biophys J 90: 2170-2178. doi: 10.1529/biophysj.105.073510
|
| [11] |
Mouritsen OG, Zuckermann MJ (2004) What's so special about cholesterol? Lipids 39: 1101-1113. doi: 10.1007/s11745-004-1336-x
|
| [12] |
Wang J, Megha, London E (2004) Relationship between sterol/steroid structure and participation in ordered lipid domains (lipid rafts): implications for lipid raft structure and function. Biochemistry 43: 1010-1018. doi: 10.1021/bi035696y
|
| [13] |
Wenz JJ, Barrantes FJ (2003) Steroid structural requirements for stabilizing or disrupting lipid domains. Biochemistry 42: 14267-14276. doi: 10.1021/bi035759c
|
| [14] |
Xu X, Bittman R, Duportail G, et al. (2001) Effect of the structure of natural sterols and sphingolipids on the formation of ordered sphingolipid/sterol domains (rafts) Comparison of cholesterol to plant, fungal, and disease-associated sterols and comparison of sphingomyelin, cerebrosides, and ceramide. J Biol Chem 276: 33540-33546. doi: 10.1074/jbc.M104776200
|
| [15] |
Xu X, London E (2000) The effect of sterol structure on membrane lipid domains reveals how cholesterol can induce lipid domain formation. Biochemistry 39: 843-849. doi: 10.1021/bi992543v
|
| [16] | Bernardino de la Serna J, Schutz GJ, Eggeling C, et al. (2016) There is no simple model of the plasma membrane organization. Front Cell Dev Biol 4: art106. |
| [17] |
van Meer G, Stelzer EH, Wijnaendts-van-Resandt RW, et al. (1987) Sorting of sphingolipids in epithelial (Madin-Darby canine kidney) cells. J Cell Biol 105: 1623-1635. doi: 10.1083/jcb.105.4.1623
|
| [18] |
Simons K, Ikonen E (1997) Functional rafts in cell membranes. Nature 387: 569-572. doi: 10.1038/42408
|
| [19] |
Gupta N, DeFranco AL (2007) Lipid rafts and B cell signaling. Semin Cell Dev Biol 18: 616-626. doi: 10.1016/j.semcdb.2007.07.009
|
| [20] |
Ikonen E (2001) Roles of lipid rafts in membrane transport. Curr Opin Cell Biol 13: 470-477. doi: 10.1016/S0955-0674(00)00238-6
|
| [21] |
Lasserre R, Guo XJ, Conchonaud F, et al. (2008) Raft nanodomains contribute to Akt/PKB plasma membrane recruitment and activation. Nat Chem Biol 4: 538-547. doi: 10.1038/nchembio.103
|
| [22] |
Orlowski S, Comera C, Terce F, et al. (2007) Lipid rafts: dream or reality for cholesterol transporters? Eur Biophys J 36: 869-885. doi: 10.1007/s00249-007-0193-8
|
| [23] |
Orlowski S, Martin S, Escargueil A (2006) P-glycoprotein and 'lipid rafts': some ambiguous mutual relationships (floating on them, building them or meeting them by chance?). Cell Mol Life Sci 63: 1038-1059. doi: 10.1007/s00018-005-5554-9
|
| [24] |
Owen DM, Magenau A, Williamson D, et al. (2012) The lipid raft hypothesis revisited—new insights on raft composition and function from super-resolution fluorescence microscopy. Bioessays 34: 739-747. doi: 10.1002/bies.201200044
|
| [25] | Simons K, Toomre D (2000) Lipid rafts and signal transduction. Nat Rev Mol Cell Biol 1: 31-39. |
| [26] |
Carquin M, D'Auria L, Pollet H, et al. (2016) Recent progress on lipid lateral heterogeneity in plasma membranes: From rafts to submicrometric domains. Prog Lipid Res 62: 1-24. doi: 10.1016/j.plipres.2015.12.004
|
| [27] |
Jacobson K, Mouritsen OG, Anderson RG (2007) Lipid rafts: at a crossroad between cell biology and physics. Nat Cell Biol 9: 7-14. doi: 10.1038/ncb0107-7
|
| [28] |
Mayor S, Rao M (2004) Rafts: scale-dependent, active lipid organization at the cell surface. Traffic 5: 231-240. doi: 10.1111/j.1600-0854.2004.00172.x
|
| [29] |
Pike LJ (2004) Lipid rafts: heterogeneity on the high seas. Biochem J 378: 281-292. doi: 10.1042/bj20031672
|
| [30] |
Simons K, Gerl MJ (2010) Revitalizing membrane rafts: new tools and insights. Nat Rev Mol Cell Biol 11: 688-699. doi: 10.1038/nrm2977
|
| [31] |
Wustner D (2007) Fluorescent sterols as tools in membrane biophysics and cell biology. Chem Phys Lipids 146: 1-25. doi: 10.1016/j.chemphyslip.2006.12.004
|
| [32] |
Baker BY, Epand RF, Epand RM, et al. (2007) Cholesterol binding does not predict activity of the steroidogenic acute regulatory protein, StAR. J Biol Chem 282: 10223-10232. doi: 10.1074/jbc.M611221200
|
| [33] |
Honigmann A, Pralle A (2016) Compartmentalization of the Cell Membrane. J Mol Biol 428: 4739-4748. doi: 10.1016/j.jmb.2016.09.022
|
| [34] |
Lenne PF, Wawrezinieck L, Conchonaud F, et al. (2006) Dynamic molecular confinement in the plasma membrane by microdomains and the cytoskeleton meshwork. EMBO J 25: 3245-3256. doi: 10.1038/sj.emboj.7601214
|
| [35] | van Zanten TS, Mayor S (2015) Current approaches to studying membrane organization. F1000Research: 6868. |
| [36] |
Lingwood D, Simons K (2010) Lipid rafts as a membrane-organizing principle. Science 327: 46-50. doi: 10.1126/science.1174621
|
| [37] |
Devaux PF, Morris R (2004) Transmembrane asymmetry and lateral domains in biological membranes. Traffic 5: 241-246. doi: 10.1111/j.1600-0854.2004.0170.x
|
| [38] |
Lorent JH, Levental I (2015) Structural determinants of protein partitioning into ordered membrane domains and lipid rafts. Chem Phys Lipids 192: 23-32. doi: 10.1016/j.chemphyslip.2015.07.022
|
| [39] |
Chum T, Glatzova D, Kvicalova Z, et al. (2016) The role of palmitoylation and transmembrane domain in sorting of transmembrane adaptor proteins. J Cell Sci 129: 95-107. doi: 10.1242/jcs.175190
|
| [40] |
Levental I, Grzybek M, Simons K (2010) Greasing their way: lipid modifications determine protein association with membrane rafts. Biochemistry 49: 6305-6316. doi: 10.1021/bi100882y
|
| [41] |
Anderson RG, Jacobson K (2002) A role for lipid shells in targeting proteins to caveolae, rafts, and other lipid domains. Science 296: 1821-1825. doi: 10.1126/science.1068886
|
| [42] |
Kaiser HJ, Orlowski A, Rog T, et al. (2011) Lateral sorting in model membranes by cholesterol-mediated hydrophobic matching. Proc Natl Acad Sci USA 108: 16628-16633. doi: 10.1073/pnas.1103742108
|
| [43] |
Nyholm TK (2015) Lipid-protein interplay and lateral organization in biomembranes. Chem Phys Lipids 189: 48-55. doi: 10.1016/j.chemphyslip.2015.05.008
|
| [44] |
Fujiwara T, Ritchie K, Murakoshi H, et al. (2002) Phospholipids undergo hop diffusion in compartmentalized cell membrane. J Cell Biol 157: 1071-1081. doi: 10.1083/jcb.200202050
|
| [45] | Braccia A, Villani M, Immerdal L, et al. (2003) Microvillar membrane microdomains exist at physiological temperature. Role of galectin-4 as lipid raft stabilizer revealed by "superrafts". J Biol Chem 278: 15679-15684. |
| [46] |
Hansen GH, Immerdal L, Thorsen E, et al. (2001) Lipid rafts exist as stable cholesterol-independent microdomains in the brush border membrane of enterocytes. J Biol Chem 276: 32338-32344. doi: 10.1074/jbc.M102667200
|
| [47] |
Danielsen EM, Hansen GH (2003) Lipid rafts in epithelial brush borders: atypical membrane microdomains with specialized functions. Biochim Biophys Acta 1617: 1-9. doi: 10.1016/j.bbamem.2003.09.005
|
| [48] |
Meder D, Moreno MJ, Verkade P, et al. (2006) Phase coexistence and connectivity in the apical membrane of polarized epithelial cells. Proc Natl Acad Sci USA 103: 329-334. doi: 10.1073/pnas.0509885103
|
| [49] |
Lingwood D, Ries J, Schwille P, et al. (2008) Plasma membranes are poised for activation of raft phase coalescence at physiological temperature. Proc Natl Acad Sci USA 105: 10005-10010. doi: 10.1073/pnas.0804374105
|
| [50] |
Mizuno H, Abe M, Dedecker P, et al. (2011) Fluorescent probes for superresolution imaging of lipid domains on the plasma membrane. Chem Sci 2: 1548-1553. doi: 10.1039/c1sc00169h
|
| [51] |
Carquin M, Conrard L, Pollet H, et al. (2015) Cholesterol segregates into submicrometric domains at the living erythrocyte membrane: evidence and regulation. Cell Mol Life Sci 72: 4633-4651. doi: 10.1007/s00018-015-1951-x
|
| [52] |
Carquin M, Pollet H, Veiga-da-Cunha M, et al. (2014) Endogenous sphingomyelin segregates into submicrometric domains in the living erythrocyte membrane. J Lipid Res 55: 1331-1342. doi: 10.1194/jlr.M048538
|
| [53] |
D'Auria L, Van der Smissen P, Bruyneel F, et al. (2011) Segregation of fluorescent membrane lipids into distinct micrometric domains: evidence for phase compartmentation of natural lipids? PLoS One 6: e17021. doi: 10.1371/journal.pone.0017021
|
| [54] |
Frisz JF, Klitzing HA, Lou K, et al. (2013) Sphingolipid domains in the plasma membranes of fibroblasts are not enriched with cholesterol. J Biol Chem 288: 16855-16861. doi: 10.1074/jbc.M113.473207
|
| [55] |
Frisz JF, Lou K, Klitzing HA, et al. (2013) Direct chemical evidence for sphingolipid domains in the plasma membranes of fibroblasts. Proc Natl Acad Sci USA 110: E613-622. doi: 10.1073/pnas.1216585110
|
| [56] |
Magenau A, Benzing C, Proschogo N, et al. (2011) Phagocytosis of IgG-coated polystyrene beads by macrophages induces and requires high membrane order. Traffic 12: 1730-1743. doi: 10.1111/j.1600-0854.2011.01272.x
|
| [57] |
Stancevic B, Kolesnick R (2010) Ceramide-rich platforms in transmembrane signaling. FEBS Lett 584: 1728-1740. doi: 10.1016/j.febslet.2010.02.026
|
| [58] |
Garcia-Ruiz C, Morales A, Fernandez-Checa JC (2015) Glycosphingolipids and cell death: one aim, many ways. Apoptosis 20: 607-620. doi: 10.1007/s10495-015-1092-6
|
| [59] |
Contreras FX, Sot J, Ruiz-Arguello MB, et al. (2004) Cholesterol modulation of sphingomyelinase activity at physiological temperatures. Chem Phys Lipids 130: 127-134. doi: 10.1016/j.chemphyslip.2004.02.003
|
| [60] |
Cheng JP, Nichols BJ (2016) Caveolae: One Function or Many? Trends Cell Biol 26: 177-189. doi: 10.1016/j.tcb.2015.10.010
|
| [61] |
Parton RG, del Pozo MA (2013) Caveolae as plasma membrane sensors, protectors and organizers. Nat Rev Mol Cell Biol 14: 98-112. doi: 10.1038/nrm3512
|
| [62] |
Parton RG, Simons K (2007) The multiple faces of caveolae. Nat Rev Mol Cell Biol 8: 185-194. doi: 10.1038/nrm2122
|
| [63] |
Shvets E, Ludwig A, Nichols BJ (2014) News from the caves: update on the structure and function of caveolae. Curr Opin Cell Biol 29: 99-106. doi: 10.1016/j.ceb.2014.04.011
|
| [64] |
Fridolfsson HN, Roth DM, Insel PA, et al. (2014) Regulation of intracellular signaling and function by caveolin. FASEB J 28: 3823-3831. doi: 10.1096/fj.14-252320
|
| [65] |
Banning A, Tomasovic A, Tikkanen R (2011) Functional aspects of membrane association of reggie/flotillin proteins. Curr Protein Pept Sci 12: 725-735. doi: 10.2174/138920311798841708
|
| [66] |
Ortegren U, Yin L, Ost A, et al. (2006) Separation and characterization of caveolae subclasses in the plasma membrane of primary adipocytes; segregation of specific proteins and functions. FEBS J 273: 3381-3392. doi: 10.1111/j.1742-4658.2006.05345.x
|
| [67] |
Sinha B, Koster D, Ruez R, et al. (2011) Cells respond to mechanical stress by rapid disassembly of caveolae. Cell 144: 402-413. doi: 10.1016/j.cell.2010.12.031
|
| [68] |
Roper K, Corbeil D, Huttner WB (2000) Retention of prominin in microvilli reveals distinct cholesterol-based lipid micro-domains in the apical plasma membrane. Nat Cell Biol 2: 582-592. doi: 10.1038/35023524
|
| [69] |
Zimmerman B, Kelly B, McMillan BJ, et al. (2016) Crystal structure of a full-length human tetraspanin reveals a cholesterol-binding pocket. Cell 167: 1041-1051. doi: 10.1016/j.cell.2016.09.056
|
| [70] |
Hemler ME (2005) Tetraspanin functions and associated microdomains. Nat Rev Mol Cell Biol 6: 801-811. doi: 10.1038/nrm1736
|
| [71] |
Yanez-Mo M, Barreiro O, Gordon-Alonso M, et al. (2009) Tetraspanin-enriched microdomains: a functional unit in cell plasma membranes. Trends Cell Biol 19: 434-446. doi: 10.1016/j.tcb.2009.06.004
|
| [72] |
Zhang XA, Huang C (2012) Tetraspanins and cell membrane tubular structures. Cell Mol Life Sci 69: 2843-2852. doi: 10.1007/s00018-012-0954-0
|
| [73] |
Hugel B, Martinez MC, Kunzelmann C, et al. (2005) Membrane microparticles: two sides of the coin. Physiology (Bethesda) 20: 22-27. doi: 10.1152/physiol.00029.2004
|
| [74] |
Raposo G, Stoorvogel W (2013) Extracellular vesicles: exosomes, microvesicles, and friends. J Cell Biol 200: 373-383. doi: 10.1083/jcb.201211138
|
| [75] |
van der Pol E, Boing AN, Harrison P, et al. (2012) Classification, functions, and clinical relevance of extracellular vesicles. Pharmacol Rev 64: 676-705. doi: 10.1124/pr.112.005983
|
| [76] | Kravets FG, Lee J, Singh B, et al. (2000) Prostasomes: current concepts. Prostate 43: 169-174. |
| [77] |
Llorente A, van Deurs B, Sandvig K (2007) Cholesterol regulates prostasome release from secretory lysosomes in PC-3 human prostate cancer cells. Eur J Cell Biol 86: 405-415. doi: 10.1016/j.ejcb.2007.05.001
|
| [78] |
Arvidson G, Ronquist G, Wikander G, et al. (1989) Human prostasome membranes exhibit very high cholesterol/phospholipid ratios yielding high molecular ordering. Biochim Biophys Acta 984: 167-173. doi: 10.1016/0005-2736(89)90212-5
|
| [79] |
Llorente A, de Marco MC, Alonso MA (2004) Caveolin-1 and MAL are located on prostasomes secreted by the prostate cancer PC-3 cell line. J Cell Sci 117: 5343-5351. doi: 10.1242/jcs.01420
|
| [80] |
Kastelowitz N, Yin H (2014) Exosomes and microvesicles: identification and targeting by particle size and lipid chemical probes. ChemBioChem 15: 923-928. doi: 10.1002/cbic.201400043
|
| [81] |
Bagnat M, Keranen S, Shevchenko A, et al. (2000) Lipid rafts function in biosynthetic delivery of proteins to the cell surface in yeast. Proc Natl Acad Sci USA 97: 3254-3259. doi: 10.1073/pnas.97.7.3254
|
| [82] |
Dupre S, Haguenauer-Tsapis R (2003) Raft partitioning of the yeast uracil permease during trafficking along the endocytic pathway. Traffic 4: 83-96. doi: 10.1034/j.1600-0854.2003.40204.x
|
| [83] |
Lauwers E, Andre B (2006) Association of yeast transporters with detergent-resistant membranes correlates with their cell-surface location. Traffic 7: 1045-1059. doi: 10.1111/j.1600-0854.2006.00445.x
|
| [84] |
Malinska K, Malinsky J, Opekarova M, et al. (2003) Visualization of protein compartmentation within the plasma membrane of living yeast cells. Mol Biol Cell 14: 4427-4436. doi: 10.1091/mbc.E03-04-0221
|
| [85] |
Malinska K, Malinsky J, Opekarova M, et al. (2004) Distribution of Can1p into stable domains reflects lateral protein segregation within the plasma membrane of living S. cerevisiae cells. J Cell Sci 117: 6031-6041. doi: 10.1242/jcs.01493
|
| [86] |
Spira F, Mueller NS, Beck G, et al. (2012) Patchwork organization of the yeast plasma membrane into numerous coexisting domains. Nat Cell Biol 14: 640-648. doi: 10.1038/ncb2487
|
| [87] | Bagnat M, Simons K (2002) Lipid rafts in protein sorting and cell polarity in budding yeast Saccharomyces cerevisiae. Biol Chem 383: 1475-1480. |
| [88] | Wachtler V, Balasubramanian MK (2006) Yeast lipid rafts?—an emerging view. Trends Cell Biol 16: 1-4. |
| [89] |
Klose C, Ejsing CS, Garcia-Saez AJ, et al. (2010) Yeast lipids can phase-separate into micrometer-scale membrane domains. J Biol Chem 285: 30224-30232. doi: 10.1074/jbc.M110.123554
|
| [90] |
Aresta-Branco F, Cordeiro AM, Marinho HS, et al. (2011) Gel domains in the plasma membrane of Saccharomyces cerevisiae: highly ordered, ergosterol-free, and sphingolipid-enriched lipid rafts. J Biol Chem 286: 5043-5054. doi: 10.1074/jbc.M110.154435
|
| [91] |
Mongrand S, Stanislas T, Bayer EM, et al. (2010) Membrane rafts in plant cells. Trends Plant Sci 15: 656-663. doi: 10.1016/j.tplants.2010.09.003
|
| [92] |
Tapken W, Murphy AS (2015) Membrane nanodomains in plants: capturing form, function, and movement. J Exp Bot 66: 1573-1586. doi: 10.1093/jxb/erv054
|
| [93] |
Malinsky J, Opekarova M, Grossmann G, et al. (2013) Membrane microdomains, rafts, and detergent-resistant membranes in plants and fungi. Annu Rev Plant Biol 64: 501-529. doi: 10.1146/annurev-arplant-050312-120103
|
| [94] |
Lopez D, Kolter R (2010) Functional microdomains in bacterial membranes. Genes Dev 24: 1893-1902. doi: 10.1101/gad.1945010
|
| [95] |
Lopez D (2015) Molecular composition of functional microdomains in bacterial membranes. Chem Phys Lipids 192: 3-11. doi: 10.1016/j.chemphyslip.2015.08.015
|
| [96] |
Saenz JP, Grosser D, Bradley AS, et al. (2015) Hopanoids as functional analogues of cholesterol in bacterial membranes. Proc Natl Acad Sci USA 112: 11971-11976. doi: 10.1073/pnas.1515607112
|
| [97] |
Huang Z, London E (2016) Cholesterol lipids and cholesterol-containing lipid rafts in bacteria. Chem Phys Lipids 199: 11-16. doi: 10.1016/j.chemphyslip.2016.03.002
|
| [98] |
Haidekker MA, Theodorakis EA (2007) Molecular rotors--fluorescent biosensors for viscosity and flow. Org Biomol Chem 5: 1669-1678. doi: 10.1039/B618415D
|
| [99] |
Demchenko AP, Mely Y, Duportail G, et al. (2009) Monitoring biophysical properties of lipid membranes by environment-sensitive fluorescent probes. Biophys J 96: 3461-3470. doi: 10.1016/j.bpj.2009.02.012
|
| [100] |
Bagatolli LA (2006) To see or not to see: lateral organization of biological membranes and fluorescence microscopy. Biochim Biophys Acta 1758: 1541-1556. doi: 10.1016/j.bbamem.2006.05.019
|
| [101] |
Gaus K, Zech T, Harder T (2006) Visualizing membrane microdomains by Laurdan 2-photon microscopy. Mol Membr Biol 23: 41-48. doi: 10.1080/09687860500466857
|
| [102] |
Gaus K, Gratton E, Kable EP, et al. (2003) Visualizing lipid structure and raft domains in living cells with two-photon microscopy. Proc Natl Acad Sci USA 100: 15554-15559. doi: 10.1073/pnas.2534386100
|
| [103] |
Sezgin E, Sadowski T, Simons K (2014) Measuring lipid packing of model and cellular membranes with environment sensitive probes. Langmuir 30: 8160-8166. doi: 10.1021/la501226v
|
| [104] |
Kucherak OA, Oncul S, Darwich Z, et al. (2010) Switchable nile red-based probe for cholesterol and lipid order at the outer leaflet of biomembranes. J Am Chem Soc 132: 4907-4916. doi: 10.1021/ja100351w
|
| [105] |
Baumgart T, Hunt G, Farkas ER, et al. (2007) Fluorescence probe partitioning between Lo/Ld phases in lipid membranes. Biochim Biophys Acta 1768: 2182-2194. doi: 10.1016/j.bbamem.2007.05.012
|
| [106] | Solanko KA, Modzel M, Solanko LM, et al. (2015) Fluorescent sterols and cholesteryl esters as probes for intracellular cholesterol transport. Lipid Insights 8: 95-114. |
| [107] |
Gimpl G, Gehrig-Burger K (2007) Cholesterol reporter molecules. Biosci Rep 27: 335-358. doi: 10.1007/s10540-007-9060-1
|
| [108] |
Fujimoto T, Hayashi M, Iwamoto M, et al. (1997) Crosslinked plasmalemmal cholesterol is sequestered to caveolae: analysis with a new cytochemical probe. J Histochem Cytochem 45: 1197-1205. doi: 10.1177/002215549704500903
|
| [109] |
Shimada Y, Maruya M, Iwashita S, et al. (2002) The C-terminal domain of perfringolysin O is an essential cholesterol-binding unit targeting to cholesterol-rich microdomains. Eur J Biochem 269: 6195-6203. doi: 10.1046/j.1432-1033.2002.03338.x
|
| [110] | Liu SL, Sheng R, Jung JH, et al. (2017) Orthogonal lipid sensors identify transbilayer asymmetry of plasma membrane cholesterol. Nat Chem Biol 13: 268-274. |
| [111] |
Koivusalo M, Alvesalo J, Virtanen JA, et al. (2004) Partitioning of pyrene-labeled phospho- and sphingolipids between ordered and disordered bilayer domains. Biophys J 86: 923-935. doi: 10.1016/S0006-3495(04)74168-5
|
| [112] |
Koivusalo M, Jansen M, Somerharju P, et al. (2007) Endocytic trafficking of sphingomyelin depends on its acyl chain length. Mol Biol Cell 18: 5113-5123. doi: 10.1091/mbc.E07-04-0330
|
| [113] |
Pagano RE, Watanabe R, Wheatley C, et al. (1999) Use of N-[5-(5,7-dimethyl boron dipyrromethene difluoride-sphingomyelin to study membrane traffic along the endocytic pathway. Chem Phys Lipids 102: 55-63. doi: 10.1016/S0009-3084(99)00075-4
|
| [114] |
Scheidt HA, Muller P, Herrmann A, et al. (2003) The potential of fluorescent and spin-labeled steroid analogs to mimic natural cholesterol. J Biol Chem 278: 45563-45569. doi: 10.1074/jbc.M303567200
|
| [115] |
Smutzer G, Crawford BF, Yeagle PL (1986) Physical properties of the fluorescent sterol probe dehydroergosterol. Biochim Biophys Acta 862: 361-371. doi: 10.1016/0005-2736(86)90239-7
|
| [116] |
Ohvo-Rekila H, Akerlund B, Slotte JP (2000) Cyclodextrin-catalyzed extraction of fluorescent sterols from monolayer membranes and small unilamellar vesicles. Chem Phys Lipids 105: 167-178. doi: 10.1016/S0009-3084(00)00122-5
|
| [117] |
Hao M, Lin SX, Karylowski OJ, et al. (2002) Vesicular and non-vesicular sterol transport in living cells The endocytic recycling compartment is a major sterol storage organelle. J Biol Chem 277: 609-617. doi: 10.1074/jbc.M108861200
|
| [118] |
Wustner D, Herrmann A, Hao M, et al. (2002) Rapid nonvesicular transport of sterol between the plasma membrane domains of polarized hepatic cells. J Biol Chem 277: 30325-30336. doi: 10.1074/jbc.M202626200
|
| [119] |
Wustner D, Mondal M, Tabas I, et al. (2005) Direct observation of rapid internalization and intracellular transport of sterol by macrophage foam cells. Traffic 6: 396-412. doi: 10.1111/j.1600-0854.2005.00285.x
|
| [120] |
Frolov A, Petrescu A, Atshaves BP, et al. (2000) High density lipoprotein-mediated cholesterol uptake and targeting to lipid droplets in intact L-cell fibroblasts A single- and multiphoton fluorescence approach. J Biol Chem 275: 12769-12780. doi: 10.1074/jbc.275.17.12769
|
| [121] | Wustner D, Mondal M, Huang A, et al. (2004) Different transport routes for high density lipoprotein and its associated free sterol in polarized hepatic cells. J Lipid Res 45: 427-437. |
| [122] |
Liu J, Chang CC, Westover EJ, et al. (2005) Investigating the allosterism of acyl-CoA:cholesterol acyltransferase (ACAT) by using various sterols: in vitro and intact cell studies. Biochem J 391: 389-397. doi: 10.1042/BJ20050428
|
| [123] |
Mukherjee S, Zha X, Tabas I, et al. (1998) Cholesterol distribution in living cells: fluorescence imaging using dehydroergosterol as a fluorescent cholesterol analog. Biophys J 75: 1915-1925. doi: 10.1016/S0006-3495(98)77632-5
|
| [124] |
Sato SB, Ishii K, Makino A, et al. (2004) Distribution and transport of cholesterol-rich membrane domains monitored by a membrane-impermeant fluorescent polyethylene glycol-derivatized cholesterol. J Biol Chem 279: 23790-23796. doi: 10.1074/jbc.M313568200
|
| [125] |
Mazeres S, Schram V, Tocanne JF, et al. (1996) 7-nitrobenz-2-oxa-1,3-diazole-4-yl-labeled phospholipids in lipid membranes: differences in fluorescence behavior. Biophys J 71: 327-335. doi: 10.1016/S0006-3495(96)79228-7
|
| [126] |
Loura LM, Fedorov A, Prieto M (2001) Exclusion of a cholesterol analog from the cholesterol-rich phase in model membranes. Biochim Biophys Acta 1511: 236-243. doi: 10.1016/S0005-2736(01)00269-3
|
| [127] | Sezgin E, Can FB, Schneider F, et al. (2016) A comparative study on fluorescent cholesterol analogs as versatile cellular reporters. J Lipid Res 57: 299-309. |
| [128] |
Gaibelet G, Allart S, Terce F, et al. (2015) Specific cellular incorporation of a pyrene-labelled cholesterol: lipoprotein-mediated delivery toward ordered intracellular membranes. PLoS One 10: e0121563. doi: 10.1371/journal.pone.0121563
|
| [129] |
Dagher G, Donne N, Klein C, et al. (2003) HDL-mediated cholesterol uptake and targeting to lipid droplets in adipocytes. J Lipid Res 44: 1811-1820. doi: 10.1194/jlr.M300267-JLR200
|
| [130] | Sparrow CP, Patel S, Baffic J, et al. (1999) A fluorescent cholesterol analog traces cholesterol absorption in hamsters and is esterified in vivo and in vitro. J Lipid Res 40: 1747-1757. |
| [131] | Wiegand V, Chang TY, Strauss JF, 3rd, et al. (2003) Transport of plasma membrane-derived cholesterol and the function of Niemann-Pick C1 Protein. FASEB J 17: 782-784. |
| [132] |
Shrivastava S, Haldar S, Gimpl G, et al. (2009) Orientation and dynamics of a novel fluorescent cholesterol analogue in membranes of varying phase. J Phys Chem B 113: 4475-4481. doi: 10.1021/jp808309u
|
| [133] |
Huang H, McIntosh AL, Atshaves BP, et al. (2010) Use of dansyl-cholestanol as a probe of cholesterol behavior in membranes of living cells. J Lipid Res 51: 1157-1172. doi: 10.1194/jlr.M003244
|
| [134] |
Li Z, Mintzer E, Bittman R (2006) First synthesis of free cholesterol-BODIPY conjugates. J Org Chem 71: 1718-1721. doi: 10.1021/jo052029x
|
| [135] |
Bergstrom F, Mikhalyov I, Hagglof P, et al. (2002) Dimers of dipyrrometheneboron difluoride (BODIPY) with light spectroscopic applications in chemistry and biology. J Am Chem Soc 124: 196-204. doi: 10.1021/ja010983f
|
| [136] |
Marks DL, Bittman R, Pagano RE (2008) Use of Bodipy-labeled sphingolipid and cholesterol analogs to examine membrane microdomains in cells. Histochem Cell Biol 130: 819-832. doi: 10.1007/s00418-008-0509-5
|
| [137] |
Ariola FS, Li Z, Cornejo C, et al. (2009) Membrane fluidity and lipid order in ternary giant unilamellar vesicles using a new bodipy-cholesterol derivative. Biophys J 96: 2696-2708. doi: 10.1016/j.bpj.2008.12.3922
|
| [138] |
Wustner D, Solanko L, Sokol E, et al. (2011) Quantitative assessment of sterol traffic in living cells by dual labeling with dehydroergosterol and BODIPY-cholesterol. Chem Phys Lipids 164: 221-235. doi: 10.1016/j.chemphyslip.2011.01.004
|
| [139] |
Solanko LM, Honigmann A, Midtiby HS, et al. (2013) Membrane orientation and lateral diffusion of BODIPY-cholesterol as a function of probe structure. Biophys J 105: 2082-2092. doi: 10.1016/j.bpj.2013.09.031
|
| [140] |
Holtta-Vuori M, Uronen RL, Repakova J, et al. (2008) BODIPY-cholesterol: a new tool to visualize sterol trafficking in living cells and organisms. Traffic 9: 1839-1849. doi: 10.1111/j.1600-0854.2008.00801.x
|
| [141] |
Mazeres S, Lagane B, Welby M, et al. (2001) Probing the lateral organization of membranes: fluorescence repercussions of pyrene probe distribution. Spectrochim Acta A Mol Biomol Spectrosc 57: 2297-2311. doi: 10.1016/S1386-1425(01)00486-3
|
| [142] |
Le Guyader L, Le Roux C, Mazeres S, et al. (2007) Changes of the membrane lipid organization characterized by means of a new cholesterol-pyrene probe. Biophys J 93: 4462-4473. doi: 10.1529/biophysj.107.112821
|
| [143] |
Lagane B, Mazeres S, Le Grimellec C, et al. (2002) Lateral distribution of cholesterol in membranes probed by means of a pyrene-labelled cholesterol: effects of acyl chain unsaturation. Biophys Chem 95: 7-22. doi: 10.1016/S0301-4622(01)00235-6
|
| [144] |
Lecompte MF, Gaibelet G, Lebrun C, et al. (2015) Cholesterol and sphingomyelin-containing model condensed lipid monolayers: heterogeneities involving ordered microdomains assessed by two cholesterol derivatives. Langmuir 31: 11921-11931. doi: 10.1021/acs.langmuir.5b02646
|
| [145] | Astarie-Dequeker C, Le Guyader L, Malaga W, et al. (2009) Phthiocerol dimycocerosates of M. tuberculosis participate in macrophage invasion by inducing changes in the organization of plasma membrane lipids. PLoS Pathog 5: e1000289. |
| [146] |
Rohrl C, Meisslitzer-Ruppitsch C, Bittman R, et al. (2012) Combined light and electron microscopy using diaminobenzidine photooxidation to monitor trafficking of lipids derived from lipoprotein particles. Curr Pharm Biotechnol 13: 331-340. doi: 10.2174/138920112799095338
|
| [147] |
Holtta-Vuori M, Sezgin E, Eggeling C, et al. (2016) Use of BODIPY-Cholesterol (TF-Chol) for Visualizing Lysosomal Cholesterol Accumulation. Traffic 17: 1054-1057. doi: 10.1111/tra.12414
|
| [148] |
Gaibelet G, Terce F, Bertrand-Michel J, et al. (2013) 21-Methylpyrenyl-cholesterol stably and specifically associates with lipoprotein peripheral hemi-membrane: a new labelling tool. Biochem Biophys Res Commun 440: 533-538. doi: 10.1016/j.bbrc.2013.09.101
|
| [149] |
Storey SM, Gallegos AM, Atshaves BP, et al. (2007) Selective cholesterol dynamics between lipoproteins and caveolae/lipid rafts. Biochemistry 46: 13891-13906. doi: 10.1021/bi700690s
|
| [150] |
Ohsaki Y, Cheng J, Suzuki M, et al. (2009) Biogenesis of cytoplasmic lipid droplets: from the lipid ester globule in the membrane to the visible structure. Biochim Biophys Acta 1791: 399-407. doi: 10.1016/j.bbalip.2008.10.002
|
| [151] |
Mukherjee S, Soe TT, Maxfield FR (1999) Endocytic sorting of lipid analogues differing solely in the chemistry of their hydrophobic tails. J Cell Biol 144: 1271-1284. doi: 10.1083/jcb.144.6.1271
|
| [152] |
Prinz WA (2007) Non-vesicular sterol transport in cells. Prog Lipid Res 46: 297-314. doi: 10.1016/j.plipres.2007.06.002
|
| 1. | Shabana Anwar, Muhammad Kamran Jamil, Amal S. Alali, Mehwish Zegham, Aisha Javed, Extremal values of the first reformulated Zagreb index for molecular trees with application to octane isomers, 2023, 9, 2473-6988, 289, 10.3934/math.2024017 | |
| 2. | Ali N. A. Koam, Ali Ahmad, Raed Qahiti, Muhammad Azeem, Waleed Hamali, Shonak Bansal, Enhanced Chemical Insights into Fullerene Structures via Modified Polynomials, 2024, 2024, 1076-2787, 10.1155/2024/9220686 | |
| 3. | Ali Ahmad, Ali N. A. Koam, Muhammad Azeem, Reverse-degree-based topological indices of fullerene cage networks, 2023, 121, 0026-8976, 10.1080/00268976.2023.2212533 | |
| 4. | Muhammad Waheed Rasheed, Abid Mahboob, Iqra Hanif, Uses of degree-based topological indices in QSPR analysis of alkaloids with poisonous and healthful nature, 2024, 12, 2296-424X, 10.3389/fphy.2024.1381887 | |
| 5. | Shriya Negi, Vijay Kumar Bhat, Face Index of Silicon Carbide Structures: An Alternative Approach, 2024, 16, 1876-990X, 5865, 10.1007/s12633-024-03119-0 | |
| 6. | Haseeb AHMAD, Muhammad AZEEM, Face-degree-based topological descriptors of germanium phosphide, 2024, 52, 18722040, 100429, 10.1016/j.cjac.2024.100429 | |
| 7. | Belman Gautham Shenoy, Raghavendra Ananthapadmanabha, Badekara Sooryanarayana, Prasanna Poojary, Vishu Kumar Mallappa, 2024, Rational Wiener Index and Rational Schultz Index of Graphs, 180, 10.3390/engproc2023059180 |


Figures(3) / Tables(1)

Gérald Gaibelet, François Tercé, Sophie Allart, Chantal Lebrun, Xavier Collet, Nadège Jamin, Stéphane Orlowski. Fluorescent probes for detecting cholesterol-rich ordered membrane microdomains: entangled relationships between structural analogies in the membrane and functional homologies in the cell[J]. AIMS Biophysics, 2017, 4(1): 121-151. doi: 10.3934/biophy.2017.1.121

| Dimension | |f12| | |f15| | |f36| |
| 1 | 24 | 48 | 7 |
| 2 | 32 | 94 | 14 |
| 3 | 40 | 152 | 23 |
| 4 | 48 | 222 | 34 |
| 5 | 56 | 304 | 47 |
| 6 | 64 | 398 | 62 |
| 7 | 72 | 504 | 79 |
| 8 | 80 | 622 | 98 |
| . | . | . | . |
| . | . | . | . |
| . | . | . | . |
| n | 8n+16 | 6n2+28n+14 | n2+4n+2 |
DownLoad:
CSV
| Dimension | |f12| | |f14| | |f17| | |f18| |
| 1 | 6 | 0 | 0 | 0 |
| 2 | 6 | 12 | 12 | 12 |
| 3 | 6 | 24 | 24 | 60 |
| 4 | 6 | 36 | 36 | 144 |
| 5 | 6 | 48 | 48 | 264 |
| 6 | 6 | 60 | 60 | 420 |
| 7 | 6 | 72 | 72 | 612 |
| 8 | 6 | 84 | 84 | 840 |
| . | . | . | . | . |
| . | . | . | . | . |
| . | . | . | . | . |
| k | 6 | 12(k−1) | 12(k−1) | 18k2−42k+24 |
DownLoad:
CSV
| Dimension m | |f15| | |f16| | |f18| | |f∞| |
| 2 | 3 | 2(n−1) | n−1 | 20n+7 |
DownLoad:
CSV
| Dimension m | |f15| | |f16| | |f17| | |f18| | |f∞| |
| 2 | 3 | 2(n−1) | 0 | n−1 | 20n+7 |
| 3 | 2 | 2n | 1 | 3(n−1) | 20n+17 |
| 4 | 2 | 2n | 3 | 5(n−1) | 20n+27 |
| 5 | 2 | 2n | 5 | 7(n−1) | 20n+37 |
| 6 | 2 | 2n | 7 | 9(n−1) | 20n+47 |
| . | . | . | . | . | . |
| . | . | . | . | . | . |
| . | . | . | . | . | . |
| m | 2 | 2n | 2m−5 | 2mn−2m−3n+3 | 20n+10m−13 |
DownLoad:
CSV
| m | |f14| | |f15| | |f16| | |f17| | |f18| | |f20| | |f35| |
| 1 | 2n+1 | 2 | 4n−2 | 0 | 0 | 2n−1 | 0 |
| 2 | 2n+2 | 2 | 8n−2 | 2 | 2n−2 | 4n−2 | 2n−1 |
| 3 | 2n+3 | 2 | 12n−2 | 4 | 4n−4 | 6n−3 | 4n−2 |
| . | . | . | . | . | . | . | . |
| . | . | . | . | . | . | . | . |
| . | . | . | . | . | . | . | . |
| m | 2n+m | 2 | 4mn−2 | 2m−2 | 2mn−2(m+n)+2 | 2mn−m | 2mn−(m+2n)+1 |
DownLoad:
CSV
| Dimension | |f12| | |f15| | |f36| |
| 1 | 24 | 48 | 7 |
| 2 | 32 | 94 | 14 |
| 3 | 40 | 152 | 23 |
| 4 | 48 | 222 | 34 |
| 5 | 56 | 304 | 47 |
| 6 | 64 | 398 | 62 |
| 7 | 72 | 504 | 79 |
| 8 | 80 | 622 | 98 |
| . | . | . | . |
| . | . | . | . |
| . | . | . | . |
| n | 8n+16 | 6n2+28n+14 | n2+4n+2 |
| Dimension | |f12| | |f14| | |f17| | |f18| |
| 1 | 6 | 0 | 0 | 0 |
| 2 | 6 | 12 | 12 | 12 |
| 3 | 6 | 24 | 24 | 60 |
| 4 | 6 | 36 | 36 | 144 |
| 5 | 6 | 48 | 48 | 264 |
| 6 | 6 | 60 | 60 | 420 |
| 7 | 6 | 72 | 72 | 612 |
| 8 | 6 | 84 | 84 | 840 |
| . | . | . | . | . |
| . | . | . | . | . |
| . | . | . | . | . |
| k | 6 | 12(k−1) | 12(k−1) | 18k2−42k+24 |
| Dimension m | |f15| | |f16| | |f18| | |f∞| |
| 2 | 3 | 2(n−1) | n−1 | 20n+7 |
| Dimension m | |f15| | |f16| | |f17| | |f18| | |f∞| |
| 2 | 3 | 2(n−1) | 0 | n−1 | 20n+7 |
| 3 | 2 | 2n | 1 | 3(n−1) | 20n+17 |
| 4 | 2 | 2n | 3 | 5(n−1) | 20n+27 |
| 5 | 2 | 2n | 5 | 7(n−1) | 20n+37 |
| 6 | 2 | 2n | 7 | 9(n−1) | 20n+47 |
| . | . | . | . | . | . |
| . | . | . | . | . | . |
| . | . | . | . | . | . |
| m | 2 | 2n | 2m−5 | 2mn−2m−3n+3 | 20n+10m−13 |
| m | |f14| | |f15| | |f16| | |f17| | |f18| | |f20| | |f35| |
| 1 | 2n+1 | 2 | 4n−2 | 0 | 0 | 2n−1 | 0 |
| 2 | 2n+2 | 2 | 8n−2 | 2 | 2n−2 | 4n−2 | 2n−1 |
| 3 | 2n+3 | 2 | 12n−2 | 4 | 4n−4 | 6n−3 | 4n−2 |
| . | . | . | . | . | . | . | . |
| . | . | . | . | . | . | . | . |
| . | . | . | . | . | . | . | . |
| m | 2n+m | 2 | 4mn−2 | 2m−2 | 2mn−2(m+n)+2 | 2mn−m | 2mn−(m+2n)+1 |
