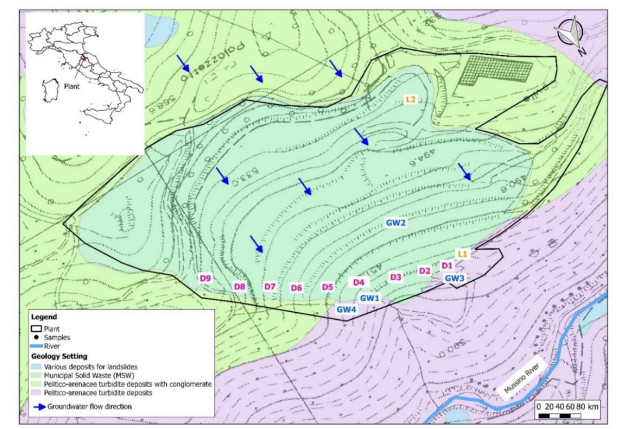
Municipal solid waste landfills leachate can cause serious environmental issues for groundwater quality. Therefore, the application of environmental tracing methods to identify groundwater contamination by municipal solid waste landfills leachate is significant. Hydrogeochemical evaluations to trace municipal solid waste landfills leachate are usually carried out. The study was carried out at a landfill in central Italy (Umbria). Samples of leachate and groundwater have been analyzed to evaluate the impact of leachates on groundwater through the comparison of their hydrogeochemical nature. Parameters like pH, Temperature (T), Electrical Conductivity (EC), redox potential (Eh) and Chemical Oxygen Demand (COD) were also measured in situ using digital instruments. Hydrogeochemical data (Na+, K+, Mg2+, Ca2+, SO42−, HCO3−, Cl−, NO3−), ionic ratios and geochemical correlations were used to confirm the processes that govern the chemistry of the spring water and to identify leachate contamination phenomena. In fact, the main geochemical diagrams (Langelier-Ludwig, Piper, Schoeller) confirm the leachate contamination in a groundwater sample. In particular, the Piper diagram shows that a sample is in Na+ – Cl- – HCO3- mixing zone, indicating a possible influence of the leachate on groundwater chemistry. As a matter of fact, some correlations between major elements, such as Cl- versus Na+ and Cl- versus HCO3-, confirm that the leachate in this study area is highly enriched in Cl- and HCO3- due to wastes dissolution and degradation processes. Further, the assessment of K+/Mg2+ ratio also confirms the presence of a sample heavily impacted from leachate contamination. These results indicate that also one basic hydrogeochemical study can be useful for fingerprinting the leachate pollution for groundwater samples.
Citation: Maurizio Barbieri, Tiziano Boschetti, Giuseppe Sappa, Francesca Andrei. Hydrogeochemistry and groundwater quality assessment in a municipal solid waste landfill (central Italy)[J]. AIMS Geosciences, 2022, 8(3): 467-487. doi: 10.3934/geosci.2022026
Municipal solid waste landfills leachate can cause serious environmental issues for groundwater quality. Therefore, the application of environmental tracing methods to identify groundwater contamination by municipal solid waste landfills leachate is significant. Hydrogeochemical evaluations to trace municipal solid waste landfills leachate are usually carried out. The study was carried out at a landfill in central Italy (Umbria). Samples of leachate and groundwater have been analyzed to evaluate the impact of leachates on groundwater through the comparison of their hydrogeochemical nature. Parameters like pH, Temperature (T), Electrical Conductivity (EC), redox potential (Eh) and Chemical Oxygen Demand (COD) were also measured in situ using digital instruments. Hydrogeochemical data (Na+, K+, Mg2+, Ca2+, SO42−, HCO3−, Cl−, NO3−), ionic ratios and geochemical correlations were used to confirm the processes that govern the chemistry of the spring water and to identify leachate contamination phenomena. In fact, the main geochemical diagrams (Langelier-Ludwig, Piper, Schoeller) confirm the leachate contamination in a groundwater sample. In particular, the Piper diagram shows that a sample is in Na+ – Cl- – HCO3- mixing zone, indicating a possible influence of the leachate on groundwater chemistry. As a matter of fact, some correlations between major elements, such as Cl- versus Na+ and Cl- versus HCO3-, confirm that the leachate in this study area is highly enriched in Cl- and HCO3- due to wastes dissolution and degradation processes. Further, the assessment of K+/Mg2+ ratio also confirms the presence of a sample heavily impacted from leachate contamination. These results indicate that also one basic hydrogeochemical study can be useful for fingerprinting the leachate pollution for groundwater samples.

| [1] |
Lee KS, Ko KS, Kim EY (2020) Application of stable isotopes and dissolved ions for monitoring landfill leachate contamination. Environ Geochem Health 42: 1387–1399. https://doi.org/10.1007/s10653-019-00427-y doi: 10.1007/s10653-019-00427-y

|
| [2] |
Mohammadzadeh H, Clark I (2008) Degradation pathways of dissolved carbon in landfill leachate traced with compound-specific 13C analysis of DOC. Isotopes Environ Health Stud 44: 267–294. https://doi.org/10.1080/10256010802309814 doi: 10.1080/10256010802309814
|
| [3] |
Mukherjee S, Mukhopadhyay S, Hashim MA, et al. (2015) Contemporary environmental issues of landfill leachate: Assessment and remedies. Crit Rev Environ Sci Technol 45: 472–590. https://doi.org/10.1080/10643389.2013.876524 doi: 10.1080/10643389.2013.876524
|
| [4] |
Qin M, Molitor H, Brazil B, et al. (2016) Recovery of nitrogen and water from landfill leachate by a microbial electrolysis cell-forward osmosis system. Bioresour Technol 200: 485–492. https://doi.org/10.1016/j.biortech.2015.10.066 doi: 10.1016/j.biortech.2015.10.066
|
| [5] |
Adeolu AO, Ada OV, Gbenga AA, et al. (2011) Assessment of groundwater contamination by leachate near a municipal solid waste landfill. Afr J Environ Sci Technol 5: 933–940. https://doi.org/10.5897/AJEST11.272 doi: 10.5897/AJEST11.272
|
| [6] |
Liu ZP, Wu WH, Shi P, et al. (2015) Characterization of dissolved organic matter in landfill leachate during the combined treatment process of air stripping, Fenton, SBR and coagulation. Waste Manag 41: 111–118. https://doi.org/10.1016/j.wasman.2015.03.044 doi: 10.1016/j.wasman.2015.03.044
|
| [7] |
Foo KY, Hameed BH (2009) An overview of landfill leachate treatment via activated carbon adsorption process. J Hazard Mater 171: 54–60. https://doi.org/10.1016/j.jhazmat.2009.06.038 doi: 10.1016/j.jhazmat.2009.06.038
|
| [8] |
De Medeiros Engelmann P, dos Santos VHJM, Barbieri CB, et al. (2018) Environmental monitoring of a landfill area through the application of carbon stable isotopes, chemical parameters and multivariate analysis. Waste Manage 76: 591–605. https://doi.org/10.1016/j.wasman.2018.02.027 doi: 10.1016/j.wasman.2018.02.027
|
| [9] |
Castañeda SS, Sucgang RJ, Almoneda RV., et al. (2012) Environmental isotopes and major ions for tracing leachate contamination from a municipal landfill in Metro Manila, Philippines. J Environ Radioact 110: 30–37. https://doi.org/10.1016/j.jenvrad.2012.01.022 doi: 10.1016/j.jenvrad.2012.01.022
|
| [10] |
Hackley LC, Liu CI, Coleman DD (1996) Environmental Isotope Characteristics of Landfill Leachates and Gases. Groundwater 34: 827–836. https://doi.org/10.1111/j.1745-6584.1996.tb02077.x doi: 10.1111/j.1745-6584.1996.tb02077.x
|
| [11] |
North JC, Frew RD, van Hale R (2006) Can stable isotopes be used to monitor landfill leachate impact on surface waters? J Geochem Explor 88: 49–53. https://doi.org/10.1016/j.gexplo.2005.08.003 doi: 10.1016/j.gexplo.2005.08.003
|
| [12] |
Naveen BP, Mahapatra DM, Sitharam TG, et al. (2017) Physico-chemical and biological characterization of urban municipal landfill leachate. Environ Pollut 220: 1–12. https://doi.org/10.1016/j.envpol.2016.09.002 doi: 10.1016/j.envpol.2016.09.002
|
| [13] |
Nigro A, Sappa G, Barbieri M (2017) Application of boron and tritium isotopes for tracing landfill contamination in groundwater. J Geochem Explor 172: 101–108. https://doi.org/10.1016/j.gexplo.2016.10.011 doi: 10.1016/j.gexplo.2016.10.011
|
| [14] |
Cozzarelli IM, Böhlke JK, Masoner J, et al. (2011) Biogeochemical evolution of a landfill leachate plume, Norman, Oklahoma. Ground Water 49: 663–687. https://doi.org/10.1111/j.1745-6584.2010.00792.x doi: 10.1111/j.1745-6584.2010.00792.x
|
| [15] |
Krishan G, Sejwal P, Bhagwat A, et al. (2021) Role of Ion Chemistry and Hydro-Geochemical Processes in Aquifer Salinization—A Case Study from a Semi-Arid Region of Haryana, India. Water 13: 617. https://doi.org/10.3390/w13050617 doi: 10.3390/w13050617
|
| [16] |
Olofsson B, Jernberg H, Rosenqvist A (2006) Tracing leachates at waste sites using geophysical and geochemical modelling. Environ Geol 49: 720–732. https://doi.org/10.1007/s00254-005-0117-9 doi: 10.1007/s00254-005-0117-9
|
| [17] |
Najet T, Belgacem A, Adel K (2019) Assessment of groundwater quality of El Ouara aquifer (southeastern Tunisia), geochemical and isotopic approaches. Arab J Geosci 12: 116. https://doi.org/10.1007/s12517-018-4203-1 doi: 10.1007/s12517-018-4203-1
|
| [18] |
Andrei F, Barbieri M, Sappa G (2021) Application of 2H and 18O isotopes for tracing municipal solid waste landfill contamination of groundwater: Two Italian case histories. Water 13: 1065. https://doi.org/10.3390/w13081065 doi: 10.3390/w13081065
|
| [19] |
Nigro A, Sappa G, Barbieri M (2018) Boron isotopes and rare earth elements in the groundwater of a landfill site. J Geochem Explor 190: 200–206. https://doi.org/10.1016/j.gexplo.2018.02.019 doi: 10.1016/j.gexplo.2018.02.019
|
| [20] |
Wimmer B, Hrad M, Huber-Humer M, et al. (2013) Stable isotope signatures for characterising the biological stability of landfilled municipal solid waste. Waste Manag 33: 2083–2090. https://doi.org/10.1016/j.wasman.2013.02.017 doi: 10.1016/j.wasman.2013.02.017
|
| [21] |
Srivastava SK, Ramanathan AL (2008) Geochemical assessment of groundwater quality in vicinity of Bhalswa landfill, Delhi, India, using graphical and multivariate statistical methods. Environ Geol 53: 1509–1528. https://doi.org/10.1007/s00254-007-0762-2 doi: 10.1007/s00254-007-0762-2
|
| [22] |
Zhang Y, Xu M, Li X, et al. (2018) Hydrochemical characteristics and multivariate statistical analysis of natural water system: A case study in Kangding County, Southwestern China. Water 10: 80. https://doi.org/10.3390/w10010080 doi: 10.3390/w10010080
|
| [23] | US. ENVIRONMENTAL PROTECTION AGENCY (2017) Low Stress (low flow) Purging and Sampling Procedure for the Collection of Groundwater Samples From Monitoring Wells. Available from: https://www.epa.gov/sites/default/files/2017-10/documents/eqasop-gw4.pdf |
| [24] | APHA AWWA WEF (2017) Standard Methods for examination of water and wastewater. 23rd Edition. American Public Health Association (APHA), the American Water Works Association (AWWA), and the Water Environment Federation (WEF). |
| [25] | APAT, CNR, IRSA (2003) 3010—Metalli e Specie Metalliche. Available from: https://www.irsa.cnr.it/wp/wp-content/uploads/2022/04/Vol1_Sez_3000_Metalli.pdf |
| [26] | APAT, CNR, IRSA (2003) 3020—Determinazione di elementi chimici mediante spettroscopia di emissione con sorgente al plasma (ICP-OES). Available from: https://www.irsa.cnr.it/wp/wp-content/uploads/2022/04/Vol1_Sez_3000_Metalli.pdf |
| [27] | APAT, CNR, IRSA (2003) 4020—Anioni (fluoruro, cloruro, nitrito, bromuro, nitrato, fosfato e solfato) in cromatografia ionica. Available from: https://www.irsa.cnr.it/wp/wp-content/uploads/2022/04/Vol2_Sez_4000_InorganiciNonMetallici.pdf |
| [28] |
Güler C, Thyne GD, McCray JE, et al. (2002) Evaluation of graphical and multivariate statistical methods for classification of water chemistry data. Hydrogeol J 10: 455–474. https://doi.org/10.1007/s10040-002-0196-6 doi: 10.1007/s10040-002-0196-6
|
| [29] |
Boschetti T (2011) Application of brine differentiation and Langelier—Ludwig plots to fresh-to-brine waters from sedimentary basins: Diagnostic potentials and limits. J Geochem Explor 108: 126–130. https://doi.org/10.1016/j.gexplo.2010.12.002 doi: 10.1016/j.gexplo.2010.12.002
|
| [30] |
Singh UK, Kumar M, Chauhan R, et al. (2008) Assessment of the impact of landfill on groundwater quality: A case study of the Pirana site in western India. Environ Monit Assess 141: 309–321. https://doi.org/10.1007/s10661-007-9897-6 doi: 10.1007/s10661-007-9897-6
|
| [31] |
Wang H, Chen Q, Wei J, et al. (2020) Geochemical Characteristics and Influencing Factors of Groundwater Fe in Seawater Intrusion Area. Water Air Soil Pollut 231: 348. https://doi.org/10.1007/s11270-020-04724-6 doi: 10.1007/s11270-020-04724-6
|









Figures(10) / Tables(3)

Maurizio Barbieri, Tiziano Boschetti, Giuseppe Sappa, Francesca Andrei. Hydrogeochemistry and groundwater quality assessment in a municipal solid waste landfill (central Italy)[J]. AIMS Geosciences, 2022, 8(3): 467-487. doi: 10.3934/geosci.2022026







 DownLoad:
DownLoad: