Citation: Xianjue Ma. Context-dependent interplay between Hippo and JNK pathway in Drosophila[J]. AIMS Genetics, 2014, 1(1): 20-33. doi: 10.3934/genet.2014.1.20

| [1] |
Pan D (2010) The hippo signaling pathway in development and cancer. Dev Cell 19: 491-505. doi: 10.1016/j.devcel.2010.09.011

|
| [2] |
Harvey KF, Zhang X, Thomas DM (2013) The Hippo pathway and human cancer. Nat Rev Cancer 13: 246-257. doi: 10.1038/nrc3458
|
| [3] |
Pan D (2007) Hippo signaling in organ size control. Genes Dev 21: 886-897. doi: 10.1101/gad.1536007
|
| [4] | Xu T, Wang W, Zhang S, et al. (1995) Identifying tumor suppressors in genetic mosaics: the Drosophila lats gene encodes a putative protein kinase. Development 121: 1053-1063. |
| [5] |
Justice RW, Zilian O, Woods DF, et al. (1995) The Drosophila tumor suppressor gene warts encodes a homolog of human myotonic dystrophy kinase and is required for the control of cell shape and proliferation. Genes Dev 9: 534-546. doi: 10.1101/gad.9.5.534
|
| [6] |
Tapon N, Harvey KF, Bell DW, et al. (2002) salvador Promotes both cell cycle exit and apoptosis in Drosophila and is mutated in human cancer cell lines. Cell 110: 467-478. doi: 10.1016/S0092-8674(02)00824-3
|
| [7] |
Kango-Singh M, Nolo R, Tao C, et al. (2002) Shar-pei mediates cell proliferation arrest during imaginal disc growth in Drosophila. Development 129: 5719-5730. doi: 10.1242/dev.00168
|
| [8] |
Harvey KF, Pfleger CM, Hariharan IK (2003) The Drosophila Mst ortholog, hippo, restricts growth and cell proliferation and promotes apoptosis. Cell 114: 457-467. doi: 10.1016/S0092-8674(03)00557-9
|
| [9] |
Wu S, Huang J, Dong J, et al. (2003) hippo encodes a Ste-20 family protein kinase that restricts cell proliferation and promotes apoptosis in conjunction with salvador and warts. Cell 114: 445-456. doi: 10.1016/S0092-8674(03)00549-X
|
| [10] |
Udan RS, Kango-Singh M, Nolo R, et al. (2003) Hippo promotes proliferation arrest and apoptosis in the Salvador/Warts pathway. Nat Cell Biol 5: 914-920. doi: 10.1038/ncb1050
|
| [11] |
Pantalacci S, Tapon N, Leopold P (2003) The Salvador partner Hippo promotes apoptosis and cell-cycle exit in Drosophila. Nat Cell Biol 5: 921-927. doi: 10.1038/ncb1051
|
| [12] |
Lai ZC, Wei X, Shimizu T, et al. (2005) Control of cell proliferation and apoptosis by mob as tumor suppressor, mats. Cell 120: 675-685. doi: 10.1016/j.cell.2004.12.036
|
| [13] |
Huang J, Wu S, Barrera J, et al. (2005) The Hippo signaling pathway coordinately regulates cell proliferation and apoptosis by inactivating Yorkie, the Drosophila Homolog of YAP. Cell 122: 421-434. doi: 10.1016/j.cell.2005.06.007
|
| [14] |
Oh H, Irvine KD (2008) In vivo regulation of Yorkie phosphorylation and localization. Development 135: 1081-1088. doi: 10.1242/dev.015255
|
| [15] |
Wu S, Liu Y, Zheng Y, et al. (2008) The TEAD/TEF family protein Scalloped mediates transcriptional output of the Hippo growth-regulatory pathway. Dev Cell 14: 388-398. doi: 10.1016/j.devcel.2008.01.007
|
| [16] |
Zhang L, Ren F, Zhang Q, et al. (2008) The TEAD/TEF family of transcription factor Scalloped mediates Hippo signaling in organ size control. Dev Cell 14: 377-387. doi: 10.1016/j.devcel.2008.01.006
|
| [17] |
Goulev Y, Fauny JD, Gonzalez-Marti B, et al. (2008) SCALLOPED interacts with YORKIE, the nuclear effector of the hippo tumor-suppressor pathway in Drosophila. Curr Biol 18: 435-441. doi: 10.1016/j.cub.2008.02.034
|
| [18] |
Staley BK, Irvine KD (2012) Hippo signaling in Drosophila: recent advances and insights. Dev Dyn 241: 3-15. doi: 10.1002/dvdy.22723
|
| [19] |
Yu FX, Guan KL (2013) The Hippo pathway: regulators and regulations. Genes Dev 27: 355-371. doi: 10.1101/gad.210773.112
|
| [20] | Mo JS, Park HW, Guan KL (2014) The Hippo signaling pathway in stem cell biology and cancer. EMBO Rep 15: 642-656. |
| [21] | Kyriakis JM, Avruch J (1990) pp54 microtubule-associated protein 2 kinase. A novel serine/threonine protein kinase regulated by phosphorylation and stimulated by poly-L-lysine. J Biol Chem 265: 17355-17363. |
| [22] |
Davis RJ (2000) Signal transduction by the JNK group of MAP kinases. Cell 103: 239-252. doi: 10.1016/S0092-8674(00)00116-1
|
| [23] |
Weston CR, Davis RJ (2007) The JNK signal transduction pathway. Curr Opin Cell Biol 19: 142-149. doi: 10.1016/j.ceb.2007.02.001
|
| [24] | Angel P, Karin M (1991) The role of Jun, Fos and the AP-1 complex in cell-proliferation and transformation. Biochim Biophys Acta 1072: 129-157. |
| [25] |
Igaki T (2009) Correcting developmental errors by apoptosis: lessons from Drosophila JNK signaling. Apoptosis 14: 1021-1028. doi: 10.1007/s10495-009-0361-7
|
| [26] |
Chen F (2012) JNK-induced apoptosis, compensatory growth, and cancer stem cells. Cancer Res 72: 379-386. doi: 10.1158/0008-5472.CAN-11-1982
|
| [27] |
Rios-Barrera LD, Riesgo-Escovar JR (2013) Regulating cell morphogenesis: the Drosophila Jun N-terminal kinase pathway. Genesis 51: 147-162. doi: 10.1002/dvg.22354
|
| [28] |
Uhlirova M, Jasper H, Bohmann D (2005) Non-cell-autonomous induction of tissue overgrowth by JNK/Ras cooperation in a Drosophila tumor model. Proc Natl Acad Sci U S A 102: 13123-13128. doi: 10.1073/pnas.0504170102
|
| [29] |
Cordero JB, Macagno JP, Stefanatos RK, et al. (2010) Oncogenic Ras diverts a host TNF tumor suppressor activity into tumor promoter. Dev Cell 18: 999-1011. doi: 10.1016/j.devcel.2010.05.014
|
| [30] |
Igaki T, Pastor-Pareja JC, Aonuma H, et al. (2009) Intrinsic tumor suppression and epithelial maintenance by endocytic activation of Eiger/TNF signaling in Drosophila. Dev Cell 16: 458-465. doi: 10.1016/j.devcel.2009.01.002
|
| [31] | Kux K, Pitsouli C (2014) Tissue communication in regenerative inflammatory signaling: lessons from the fly gut. Front Cell Infect Microbiol 4: 49. |
| [32] |
Amcheslavsky A, Jiang J, Ip YT (2009) Tissue damage-induced intestinal stem cell division in Drosophila. Cell Stem Cell 4: 49-61. doi: 10.1016/j.stem.2008.10.016
|
| [33] |
Apidianakis Y, Rahme LG (2011) Drosophila melanogaster as a model for human intestinal infection and pathology. Dis Model Mech 4: 21-30. doi: 10.1242/dmm.003970
|
| [34] |
Ohlstein B, Spradling A (2007) Multipotent Drosophila intestinal stem cells specify daughter cell fates by differential notch signaling. Science 315: 988-992. doi: 10.1126/science.1136606
|
| [35] |
Lucchetta EM, Ohlstein B (2012) The Drosophila midgut: a model for stem cell driven tissue regeneration. Wiley Interdiscip Rev Dev Biol 1: 781-788. doi: 10.1002/wdev.51
|
| [36] |
Staley BK, Irvine KD (2010) Warts and Yorkie mediate intestinal regeneration by influencing stem cell proliferation. Curr Biol 20: 1580-1587. doi: 10.1016/j.cub.2010.07.041
|
| [37] |
Buchon N, Broderick NA, Chakrabarti S, et al. (2009) Invasive and indigenous microbiota impact intestinal stem cell activity through multiple pathways in Drosophila. Genes Dev 23: 2333-2344. doi: 10.1101/gad.1827009
|
| [38] |
Jiang H, Patel PH, Kohlmaier A, et al. (2009) Cytokine/Jak/Stat signaling mediates regeneration and homeostasis in the Drosophila midgut. Cell 137: 1343-1355. doi: 10.1016/j.cell.2009.05.014
|
| [39] |
Biteau B, Hochmuth CE, Jasper H (2008) JNK activity in somatic stem cells causes loss of tissue homeostasis in the aging Drosophila gut. Cell Stem Cell 3: 442-455. doi: 10.1016/j.stem.2008.07.024
|
| [40] |
Ren F, Wang B, Yue T, et al. (2010) Hippo signaling regulates Drosophila intestine stem cell proliferation through multiple pathways. Proc Natl Acad Sci U S A 107: 21064-21069. doi: 10.1073/pnas.1012759107
|
| [41] |
Shaw RL, Kohlmaier A, Polesello C, et al. (2010) The Hippo pathway regulates intestinal stem cell proliferation during Drosophila adult midgut regeneration. Development 137: 4147-4158. doi: 10.1242/dev.052506
|
| [42] |
Cordero JB, Stefanatos RK, Scopelliti A, et al. (2012) Inducible progenitor-derived Wingless regulates adult midgut regeneration in Drosophila. EMBO J 31: 3901-3917. doi: 10.1038/emboj.2012.248
|
| [43] |
Karpowicz P, Perez J, Perrimon N (2010) The Hippo tumor suppressor pathway regulates intestinal stem cell regeneration. Development 137: 4135-4145. doi: 10.1242/dev.060483
|
| [44] |
Biteau B, Jasper H (2011) EGF signaling regulates the proliferation of intestinal stem cells in Drosophila. Development 138: 1045-1055. doi: 10.1242/dev.056671
|
| [45] |
Myant KB, Scopelliti A, Haque S, et al. (2013) Rac1 drives intestinal stem cell proliferation and regeneration. Cell Cycle 12: 2973-2977. doi: 10.4161/cc.26031
|
| [46] |
Ren F, Shi Q, Chen Y, et al. (2013) Drosophila Myc integrates multiple signaling pathways to regulate intestinal stem cell proliferation during midgut regeneration. Cell Res 23: 1133-1146. doi: 10.1038/cr.2013.101
|
| [47] |
Worley MI, Setiawan L, Hariharan IK (2012) Regeneration and transdetermination in Drosophila imaginal discs. Annu Rev Genet 46: 289-310. doi: 10.1146/annurev-genet-110711-155637
|
| [48] |
Bergantinos C, Corominas M, Serras F (2010) Cell death-induced regeneration in wing imaginal discs requires JNK signalling. Development 137: 1169-1179. doi: 10.1242/dev.045559
|
| [49] |
Bosch M, Serras F, Martin-Blanco E, et al. (2005) JNK signaling pathway required for wound healing in regenerating Drosophila wing imaginal discs. Dev Biol 280: 73-86. doi: 10.1016/j.ydbio.2005.01.002
|
| [50] |
Ryoo HD, Gorenc T, Steller H (2004) Apoptotic cells can induce compensatory cell proliferation through the JNK and the Wingless signaling pathways. Dev Cell 7: 491-501. doi: 10.1016/j.devcel.2004.08.019
|
| [51] |
Grusche FA, Degoutin JL, Richardson HE, et al. (2011) The Salvador/Warts/Hippo pathway controls regenerative tissue growth in Drosophila melanogaster. Dev Biol 350: 255-266. doi: 10.1016/j.ydbio.2010.11.020
|
| [52] |
Sun G, Irvine KD (2011) Regulation of Hippo signaling by Jun kinase signaling during compensatory cell proliferation and regeneration, and in neoplastic tumors. Dev Biol 350: 139-151. doi: 10.1016/j.ydbio.2010.11.036
|
| [53] |
Wu M, Pastor-Pareja JC, Xu T (2010) Interaction between Ras(V12) and scribbled clones induces tumour growth and invasion. Nature 463: 545-548. doi: 10.1038/nature08702
|
| [54] |
Smith-Bolton RK, Worley MI, Kanda H, et al. (2009) Regenerative growth in Drosophila imaginal discs is regulated by Wingless and Myc. Dev Cell 16: 797-809. doi: 10.1016/j.devcel.2009.04.015
|
| [55] |
Morata G, Ripoll P (1975) Minutes: mutants of drosophila autonomously affecting cell division rate. Dev Biol 42: 211-221. doi: 10.1016/0012-1606(75)90330-9
|
| [56] |
Levayer R, Moreno E (2013) Mechanisms of cell competition: themes and variations. J Cell Biol 200: 689-698. doi: 10.1083/jcb.201301051
|
| [57] |
Vincent JP, Fletcher AG, Baena-Lopez LA (2013) Mechanisms and mechanics of cell competition in epithelia. Nat Rev Mol Cell Biol 14: 581-591. doi: 10.1038/nrm3639
|
| [58] |
Tyler DM, Li W, Zhuo N, et al. (2007) Genes affecting cell competition in Drosophila. Genetics 175: 643-657. doi: 10.1534/genetics.106.061929
|
| [59] |
Moreno E, Basler K, Morata G (2002) Cells compete for decapentaplegic survival factor to prevent apoptosis in Drosophila wing development. Nature 416: 755-759. doi: 10.1038/416755a
|
| [60] |
Menendez J, Perez-Garijo A, Calleja M, et al. (2010) A tumor-suppressing mechanism in Drosophila involving cell competition and the Hippo pathway. Proc Natl Acad Sci U S A 107: 14651-14656. doi: 10.1073/pnas.1009376107
|
| [61] | Grzeschik NA, Parsons LM, Richardson HE (2010) Lgl, the SWH pathway and tumorigenesis: It's a matter of context & competition! Cell Cycle 9: 3202-3212. |
| [62] |
Chen CL, Schroeder MC, Kango-Singh M, et al. (2012) Tumor suppression by cell competition through regulation of the Hippo pathway. Proc Natl Acad Sci U S A 109: 484-489. doi: 10.1073/pnas.1113882109
|
| [63] |
Enomoto M, Igaki T (2013) Src controls tumorigenesis via JNK-dependent regulation of the Hippo pathway in Drosophila. EMBO Rep 14: 65-72. doi: 10.1038/embor.2012.185
|
| [64] |
Brumby AM, Richardson HE (2003) scribble mutants cooperate with oncogenic Ras or Notch to cause neoplastic overgrowth in Drosophila. EMBO J 22: 5769-5779. doi: 10.1093/emboj/cdg548
|
| [65] |
Tamori Y, Bialucha CU, Tian AG, et al. (2010) Involvement of Lgl and Mahjong/VprBP in cell competition. PLoS Biol 8: e1000422. doi: 10.1371/journal.pbio.1000422
|
| [66] |
Ohsawa S, Sugimura K, Takino K, et al. (2011) Elimination of oncogenic neighbors by JNK-mediated engulfment in Drosophila. Dev Cell 20: 315-328. doi: 10.1016/j.devcel.2011.02.007
|
| [67] |
Froldi F, Ziosi M, Garoia F, et al. (2010) The lethal giant larvae tumour suppressor mutation requires dMyc oncoprotein to promote clonal malignancy. BMC Biol 8: 33. doi: 10.1186/1741-7007-8-33
|
| [68] |
Ziosi M, Baena-Lopez LA, Grifoni D, et al. (2010) dMyc Functions Downstream of Yorkie to Promote the Supercompetitive Behavior of Hippo Pathway Mutant Cells. PLoS Genet 6: e1001140. doi: 10.1371/journal.pgen.1001140
|
| [69] |
Neto-Silva RM, de Beco S, Johnston LA (2010) Evidence for a Growth-Stabilizing Regulatory Feedback Mechanism between Myc and Yorkie, the Drosophila Homolog of Yap. Dev Cell 19: 507-520. doi: 10.1016/j.devcel.2010.09.009
|
| [70] |
Moreno E, Basler K (2004) dMyc transforms cells into super-competitors. Cell 117: 117-129. doi: 10.1016/S0092-8674(04)00262-4
|
| [71] | Grifoni D, Bellosta P (2014) Drosophila Myc: A master regulator of cellular performance. Biochim Biophys Acta. |
| [72] |
Doggett K, Grusche FA, Richardson HE, et al. (2011) Loss of the Drosophila cell polarity regulator Scribbled promotes epithelial tissue overgrowth and cooperation with oncogenic Ras-Raf through impaired Hippo pathway signaling. BMC Dev Biol 11: 57. doi: 10.1186/1471-213X-11-57
|
| [73] |
Leong GR, Goulding KR, Amin N, et al. (2009) Scribble mutants promote aPKC and JNK-dependent epithelial neoplasia independently of Crumbs. BMC Biol 7: 62. doi: 10.1186/1741-7007-7-62
|
| [74] |
Rhiner C, Lopez-Gay JM, Soldini D, et al. (2010) Flower forms an extracellular code that reveals the fitness of a cell to its neighbors in Drosophila. Dev Cell 18: 985-998. doi: 10.1016/j.devcel.2010.05.010
|
| [75] |
Grzeschik NA, Parsons LM, Allott ML, et al. (2010) Lgl, aPKC, and Crumbs regulate the Salvador/Warts/Hippo pathway through two distinct mechanisms. Curr Biol 20: 573-581. doi: 10.1016/j.cub.2010.01.055
|
| [76] |
Ballesteros-Arias L, Saavedra V, Morata G (2014) Cell competition may function either as tumour-suppressing or as tumour-stimulating factor in Drosophila. Oncogene 33: 4377-4384. doi: 10.1038/onc.2013.407
|
| [77] |
Ohsawa S, Sato Y, Enomoto M, et al. (2012) Mitochondrial defect drives non-autonomous tumour progression through Hippo signalling in Drosophila. Nature 490: 547-551. doi: 10.1038/nature11452
|
| [78] |
Igaki T, Pagliarini RA, Xu T (2006) Loss of cell polarity drives tumor growth and invasion through JNK activation in Drosophila. Curr Biol 16: 1139-1146. doi: 10.1016/j.cub.2006.04.042
|
| [79] |
Pagliarini RA, Xu T (2003) A genetic screen in Drosophila for metastatic behavior. Science 302: 1227-1231. doi: 10.1126/science.1088474
|
| [80] |
Rodrigues AB, Zoranovic T, Ayala-Camargo A, et al. (2012) Activated STAT regulates growth and induces competitive interactions independently of Myc, Yorkie, Wingless and ribosome biogenesis. Development 139: 4051-4061. doi: 10.1242/dev.076760
|
| [81] |
Brumby AM, Goulding KR, Schlosser T, et al. (2011) Identification of novel Ras-cooperating oncogenes in Drosophila melanogaster: a RhoGEF/Rho-family/JNK pathway is a central driver of tumorigenesis. Genetics 188: 105-125. doi: 10.1534/genetics.111.127910
|
| [82] |
Khoo P, Allan K, Willoughby L, et al. (2013) In Drosophila, RhoGEF2 cooperates with activated Ras in tumorigenesis through a pathway involving Rho1-Rok-Myosin-II and JNK signalling. Dis Model Mech 6: 661-678. doi: 10.1242/dmm.010066
|
| [83] | Sun G, Irvine KD (2013) Ajuba family proteins link JNK to Hippo signaling. Sci Signal 6: ra81. |
| [84] | Das Thakur M, Feng Y, Jagannathan R, et al. (2010) Ajuba LIM proteins are negative regulators of the Hippo signaling pathway. Curr Biol 20: 657-662. |
| [85] |
Uhlirova M, Bohmann D (2006) JNK- and Fos-regulated Mmp1 expression cooperates with Ras to induce invasive tumors in Drosophila. EMBO J 25: 5294-5304. doi: 10.1038/sj.emboj.7601401
|
| [86] |
Miles WO, Dyson NJ, Walker JA (2011) Modeling tumor invasion and metastasis in Drosophila. Dis Model Mech 4: 753-761. doi: 10.1242/dmm.006908
|
| [87] |
Willoughby LF, Schlosser T, Manning SA, et al. (2013) An in vivo large-scale chemical screening platform using Drosophila for anti-cancer drug discovery. Dis Model Mech 6: 521-529. doi: 10.1242/dmm.009985
|
| [88] |
Pastor-Pareja JC, Xu T (2013) Dissecting Social Cell Biology and Tumors Using Drosophila Genetics. Annu Rev Genet 47: 51-74. doi: 10.1146/annurev-genet-110711-155414
|
| [89] |
Friedl P, Alexander S (2011) Cancer invasion and the microenvironment: plasticity and reciprocity. Cell 147: 992-1009. doi: 10.1016/j.cell.2011.11.016
|
| [90] |
Ma X, Shao Y, Zheng H, et al. (2013) Src42A modulates tumor invasion and cell death via Ben/dUev1a-mediated JNK activation in Drosophila. Cell Death Dis 4: e864. doi: 10.1038/cddis.2013.392
|
| [91] |
Ma X, Yang L, Yang Y, et al. (2013) dUev1a modulates TNF-JNK mediated tumor progression and cell death in Drosophila. Dev Biol 380: 211-221. doi: 10.1016/j.ydbio.2013.05.013
|
| [92] |
Ma X, Li W, Yu H, et al. (2014) Bendless modulates JNK-mediated cell death and migration in Drosophila. Cell Death Differ 21: 407-415. doi: 10.1038/cdd.2013.154
|
| [93] |
Lamar JM, Stern P, Liu H, et al. (2012) The Hippo pathway target, YAP, promotes metastasis through its TEAD-interaction domain. Proc Natl Acad Sci U S A 109: E2441-2450. doi: 10.1073/pnas.1212021109
|
| [94] |
Vidal M, Larson DE, Cagan RL (2006) Csk-deficient boundary cells are eliminated from normal Drosophila epithelia by exclusion, migration, and apoptosis. Dev Cell 10: 33-44. doi: 10.1016/j.devcel.2005.11.007
|
| [95] | Herranz H, Hong X, Cohen SM (2012) Mutual repression by bantam miRNA and Capicua links the EGFR/MAPK and Hippo pathways in growth control. Curr Biol 22: 651-657. |
| [96] |
Srivastava A, Pastor-Pareja JC, Igaki T, et al. (2007) Basement membrane remodeling is essential for Drosophila disc eversion and tumor invasion. Proc Natl Acad Sci U S A 104: 2721-2726. doi: 10.1073/pnas.0611666104
|
| [97] |
Yuan M, Tomlinson V, Lara R, et al. (2008) Yes-associated protein (YAP) functions as a tumor suppressor in breast. Cell Death Differ 15: 1752-1759. doi: 10.1038/cdd.2008.108
|
| [98] |
Lucas EP, Khanal I, Gaspar P, et al. (2013) The Hippo pathway polarizes the actin cytoskeleton during collective migration of Drosophila border cells. J Cell Biol 201: 875-885. doi: 10.1083/jcb.201210073
|
| [99] |
Lin TH, Yeh TH, Wang TW, et al. (2014) The Hippo Pathway Controls Border Cell Migration Through Distinct Mechanisms in Outer Border Cells and Polar Cells of the Drosophila Ovary. Genetics 198: 1087-1099. doi: 10.1534/genetics.114.167346
|
| [100] |
Llense F, Martin-Blanco E (2008) JNK signaling controls border cell cluster integrity and collective cell migration. Curr Biol 18: 538-544. doi: 10.1016/j.cub.2008.03.029
|
| [101] |
Kulshammer E, Uhlirova M (2013) The actin cross-linker Filamin/Cheerio mediates tumor malignancy downstream of JNK signaling. J Cell Sci 126: 927-938. doi: 10.1242/jcs.114462
|
| [102] |
Rauskolb C, Sun S, Sun G, et al. (2014) Cytoskeletal tension inhibits Hippo signaling through an Ajuba-Warts complex. Cell 158: 143-156. doi: 10.1016/j.cell.2014.05.035
|
| [103] |
Fernandez BG, Jezowska B, Janody F (2014) Drosophila actin-Capping Protein limits JNK activation by the Src proto-oncogene. Oncogene 33: 2027-2039. doi: 10.1038/onc.2013.155
|
| [104] |
Wagner EF, Nebreda AR (2009) Signal integration by JNK and p38 MAPK pathways in cancer development. Nat Rev Cancer 9: 537-549. doi: 10.1038/nrc2694
|
| [105] |
Park HW, Guan KL (2013) Regulation of the Hippo pathway and implications for anticancer drug development. Trends Pharmacol Sci 34: 581-589. doi: 10.1016/j.tips.2013.08.006
|
| [106] |
Bubici C, Papa S (2014) JNK signalling in cancer: in need of new, smarter therapeutic targets. Br J Pharmacol 171: 24-37. doi: 10.1111/bph.12432
|
| [107] |
Gladstone M, Su TT (2011) Chemical genetics and drug screening in Drosophila cancer models. J Genet Genomics 38: 497-504. doi: 10.1016/j.jgg.2011.09.003
|
| [108] |
Gonzalez C (2013) Drosophila melanogaster: a model and a tool to investigate malignancy and identify new therapeutics. Nat Rev Cancer 13: 172-183. doi: 10.1038/nrc3461
|
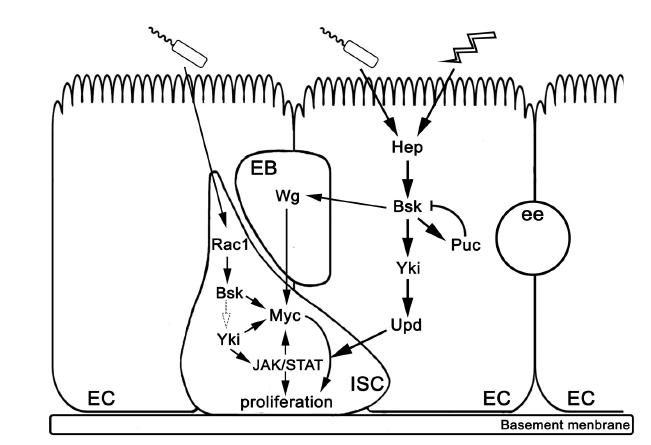
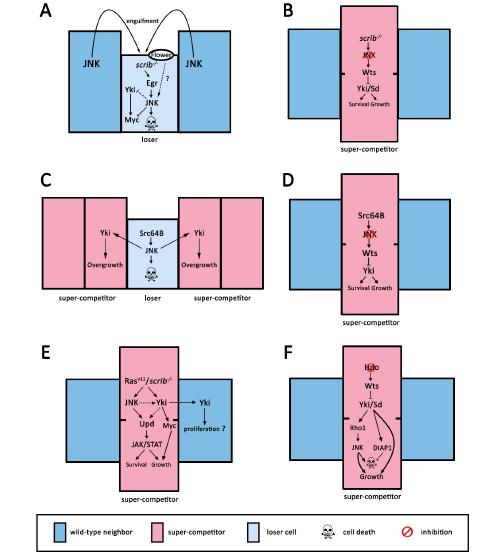
Figures(2)

Xianjue Ma. Context-dependent interplay between Hippo and JNK pathway in Drosophila[J]. AIMS Genetics, 2014, 1(1): 20-33. doi: 10.3934/genet.2014.1.20







 DownLoad:
DownLoad: