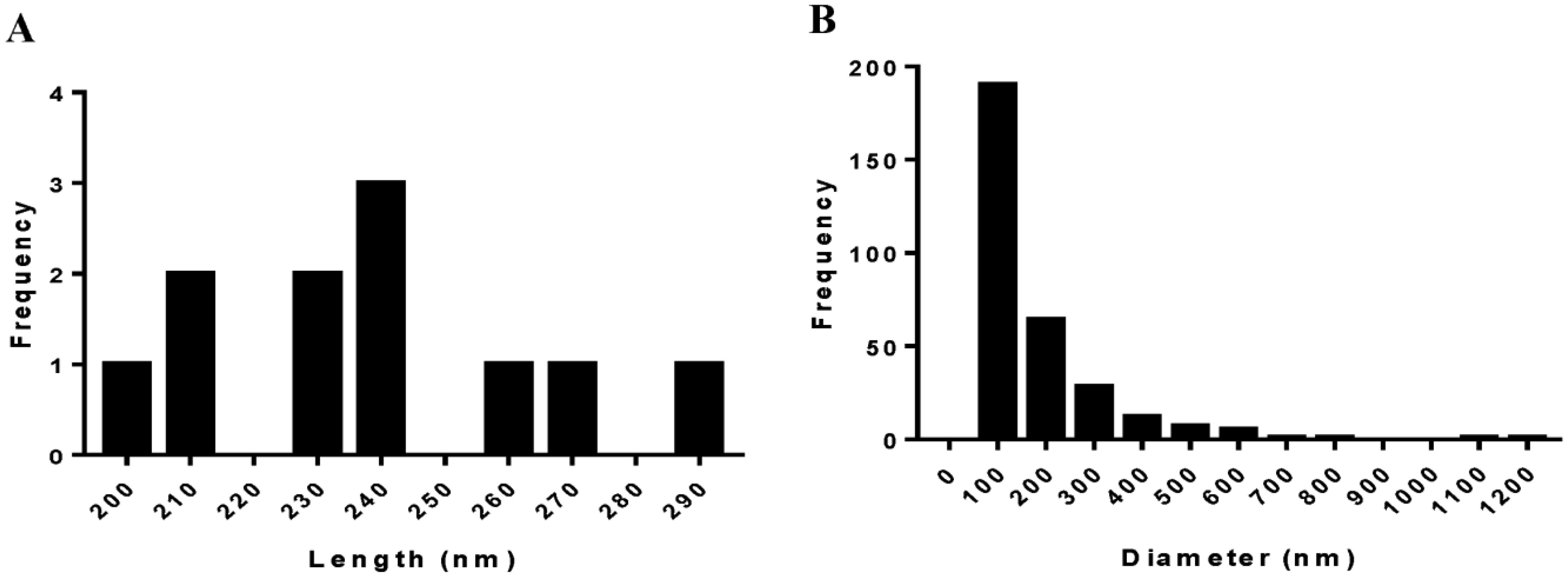
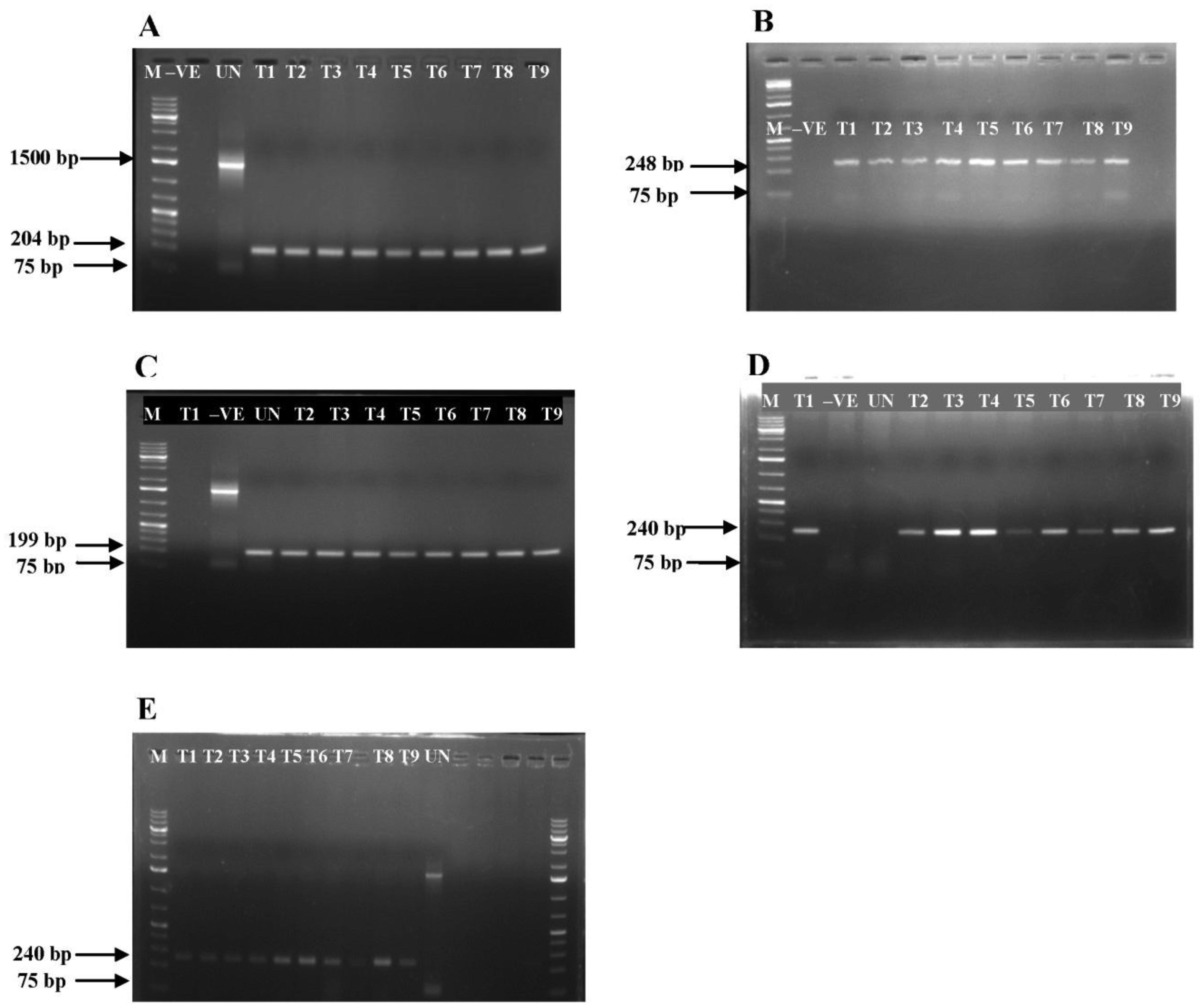
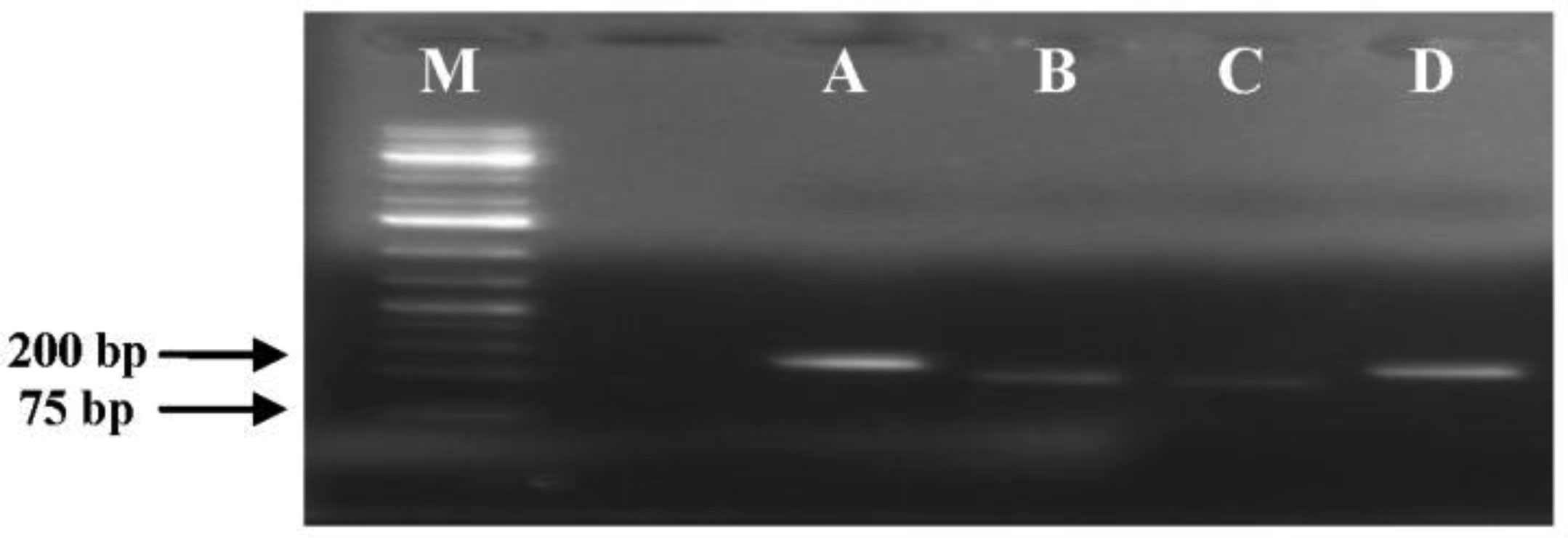
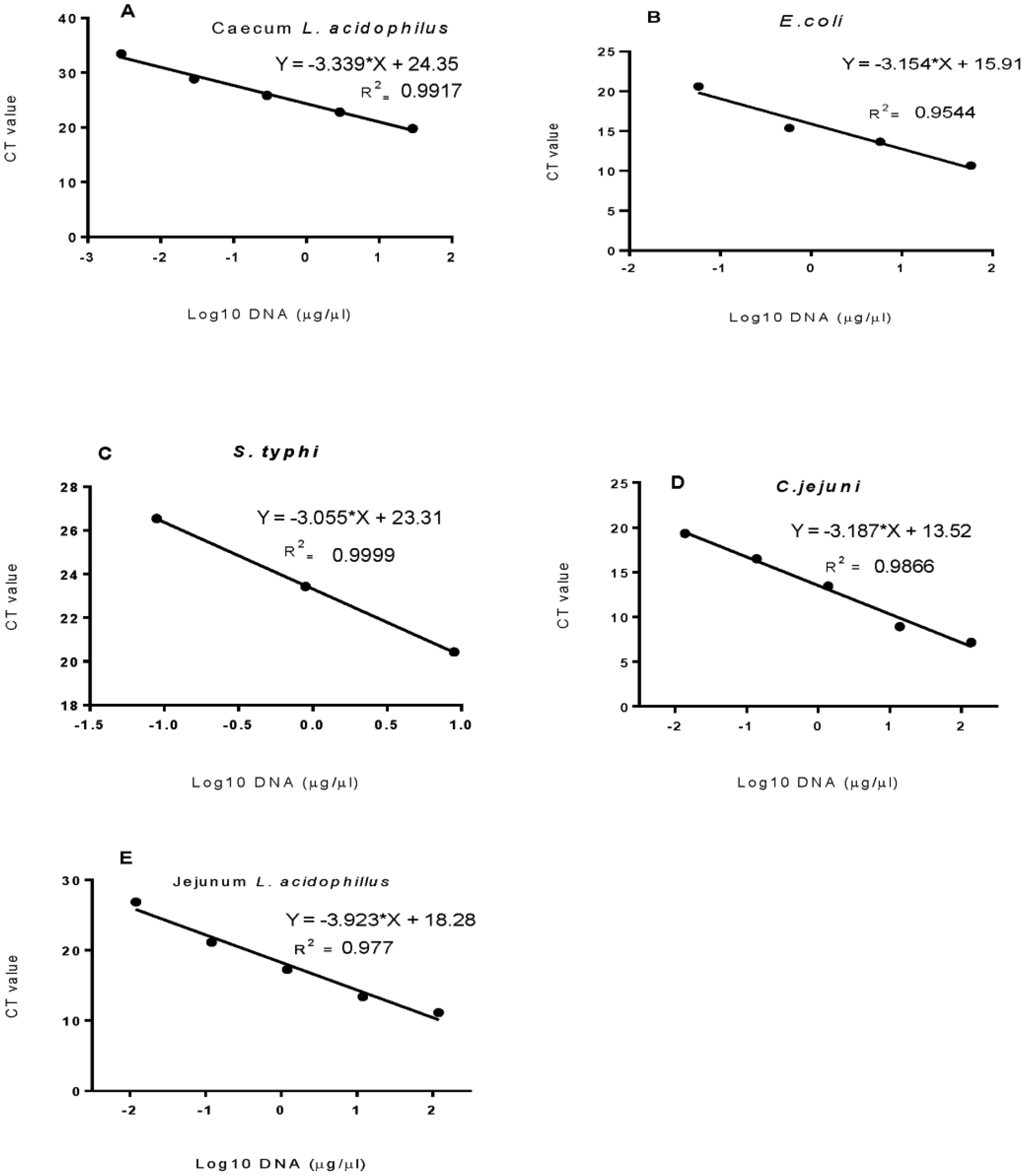
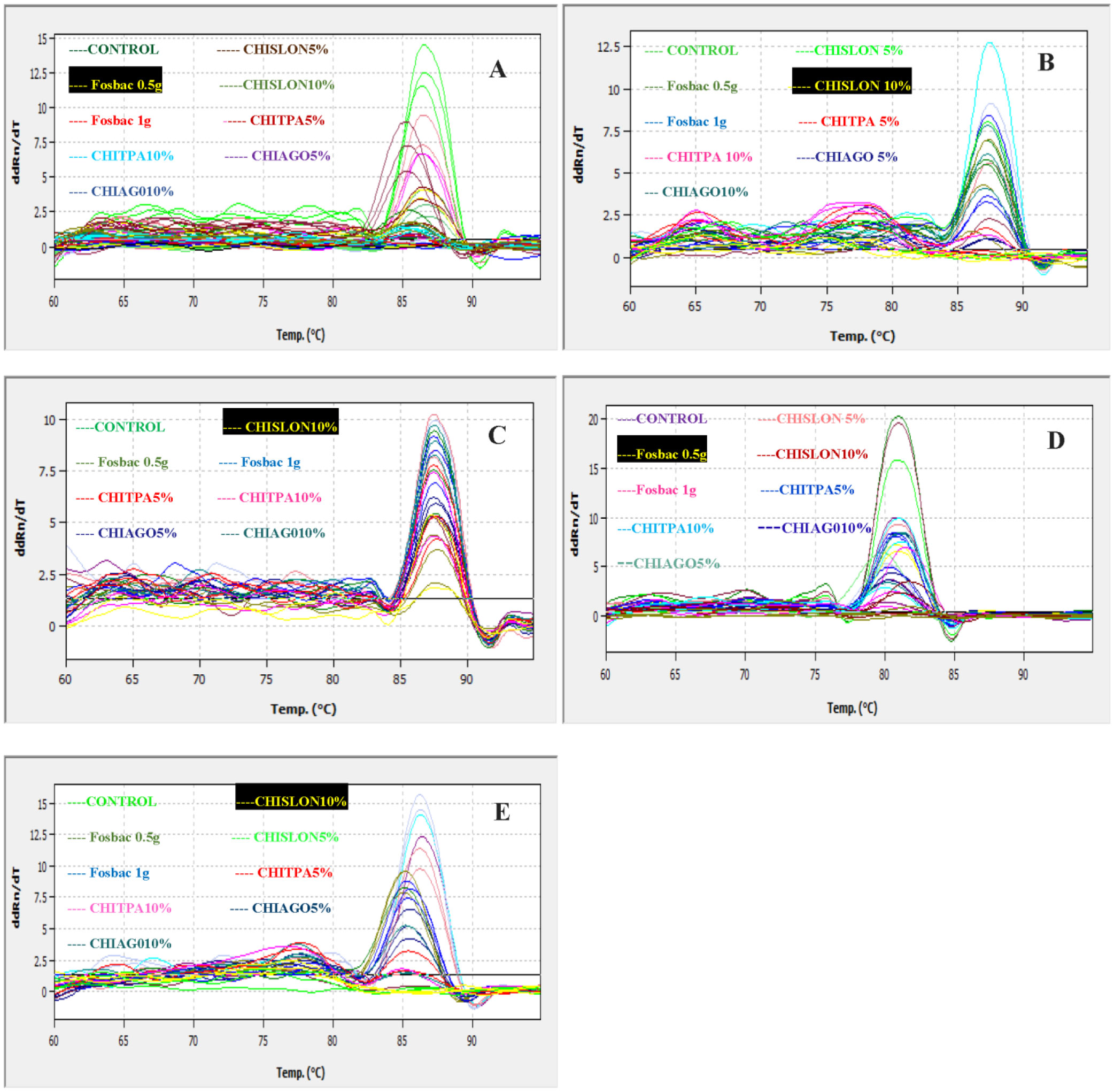
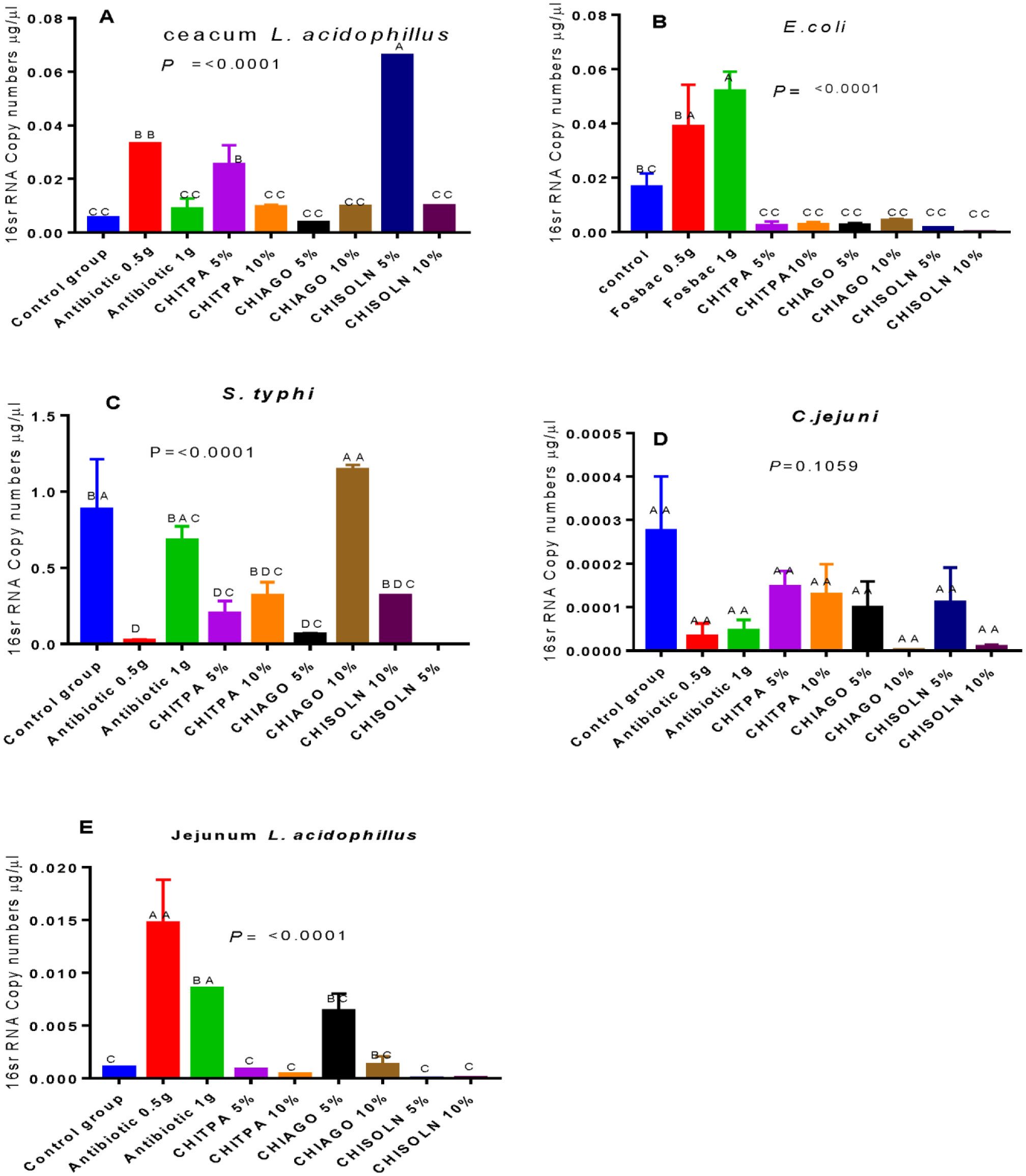
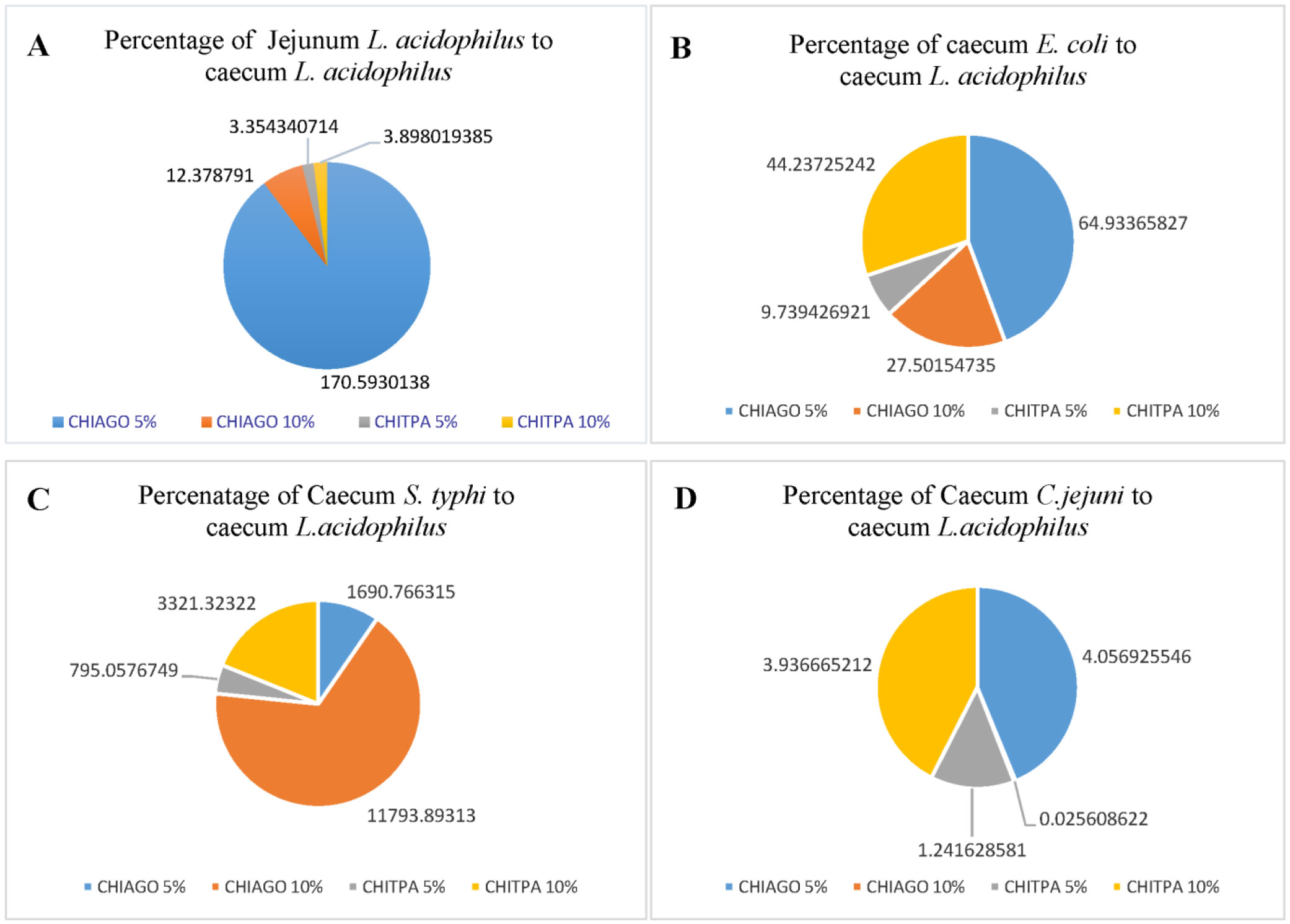
Bacterial microbes play a vital role in the nutrition and health of birds. Food passaging through the gastrointestinal tract of poultry favored microflora that have rapid growth and can adhering to the mucosal wall. The caeca are ideal habitats for diverse bacteria with effect on the host nutrition and health. Antibiotics in poultry for therapeutic and as growth promotor can decrease the number of most susceptible bacterial communities and enhance the growth of resistant bacteria. The aim of the study was to determine the effect of garlic and onion extract chitosan nanoparticles on the intestinal microflora of Rainbow Rooster Indigenous Chicken in Kenya in which a total of 18 chickens were used with 2 chickens drawn from each of the 9 groups and both caecum and jejunum content sampled with a total of 36 samples. The chickens were treated with Chitosan and Aqueous extracts of Garlic and Onion (CHIAGO), Chitosan with total Phenol, Ajoene rich extract (CHITPA) nanoparticles, and Chitosan Solution (CHISOLN) all at 5% and 10% and 1 g and 0.5 g Fosbac (Antibiotic) were applied orally to the chickens twice a week for 8 weeks. About 1.5 g of caecum and jejunum contents for micro-organisms Escherichia coli (E. coli), Salmonella typhi (S. typhi), Campylobacter jejuni (C. jejuni), Lactobacillus acidophilus (L. acidophilus), Bifidobactreum bifidium (B. bifidium) from caecum and L. acidophilus from jejunum were analyzed at 8th week of the treatment using conventional PCR to optimize bacteria 16S rRNA gene specific bands and qPCR for the 16S rRNA gene copy numbers was determined. Lactobacillus acidophilus, E. coli , S. typhi, C. jejuni from caecum and L. acidophilus from jejunum indicated specific bands in 1.2% agarose gel. The qPCR revealed primers efficiency in most of the assay with exception of the jejunum L. acidophilus assay. There was a significant differences among the treatments for L. acidophilus (p < 0.0001), E. coli (p < 0.0001), S. typhi (p < 0.0001), C. jejuni (p < 0.1059) in caecum and jejunum L. acidophilus at (p ≤ 0.0001) for 16S rRNA gene copy numbers µg/µl DNA in 1.5 g of caecum and jejunum content. The results indicated normal percentage for caecum L. acidophilus to jejunium L. acidophilus at 96.65–87.63%, 90.27% to 35% was shown in L. acidophilus to E. coli, L. acidophilus to C. jejuni was 99.97% to 95.94% and low percentage of L. acidophilus to S. typhi 16S rRNA gene copy numbers after the treatment with CHITPA and CHIAGO. Garlic and onion extract chitosan nanoparticles prepared revealed the presence of selected commensal bacteria and acceptable percentage for caecum L. acidophilus to E. coli and C. jejuni in intestine of Rainbow Rooster Chicken.
Citation: Viviana Imbua Levi Enoka, Gideon Mutie Kikuvi, Perpetual Wangui Ndung'u. Effect of garlic and onion extract chitosan nanoparticles on selected intestinal bacterial flora in indigenous rainbow rooster chicken in Kenya[J]. AIMS Molecular Science, 2021, 8(2): 98-116. doi: 10.3934/molsci.2021008
Bacterial microbes play a vital role in the nutrition and health of birds. Food passaging through the gastrointestinal tract of poultry favored microflora that have rapid growth and can adhering to the mucosal wall. The caeca are ideal habitats for diverse bacteria with effect on the host nutrition and health. Antibiotics in poultry for therapeutic and as growth promotor can decrease the number of most susceptible bacterial communities and enhance the growth of resistant bacteria. The aim of the study was to determine the effect of garlic and onion extract chitosan nanoparticles on the intestinal microflora of Rainbow Rooster Indigenous Chicken in Kenya in which a total of 18 chickens were used with 2 chickens drawn from each of the 9 groups and both caecum and jejunum content sampled with a total of 36 samples. The chickens were treated with Chitosan and Aqueous extracts of Garlic and Onion (CHIAGO), Chitosan with total Phenol, Ajoene rich extract (CHITPA) nanoparticles, and Chitosan Solution (CHISOLN) all at 5% and 10% and 1 g and 0.5 g Fosbac (Antibiotic) were applied orally to the chickens twice a week for 8 weeks. About 1.5 g of caecum and jejunum contents for micro-organisms Escherichia coli (E. coli), Salmonella typhi (S. typhi), Campylobacter jejuni (C. jejuni), Lactobacillus acidophilus (L. acidophilus), Bifidobactreum bifidium (B. bifidium) from caecum and L. acidophilus from jejunum were analyzed at 8th week of the treatment using conventional PCR to optimize bacteria 16S rRNA gene specific bands and qPCR for the 16S rRNA gene copy numbers was determined. Lactobacillus acidophilus, E. coli , S. typhi, C. jejuni from caecum and L. acidophilus from jejunum indicated specific bands in 1.2% agarose gel. The qPCR revealed primers efficiency in most of the assay with exception of the jejunum L. acidophilus assay. There was a significant differences among the treatments for L. acidophilus (p < 0.0001), E. coli (p < 0.0001), S. typhi (p < 0.0001), C. jejuni (p < 0.1059) in caecum and jejunum L. acidophilus at (p ≤ 0.0001) for 16S rRNA gene copy numbers µg/µl DNA in 1.5 g of caecum and jejunum content. The results indicated normal percentage for caecum L. acidophilus to jejunium L. acidophilus at 96.65–87.63%, 90.27% to 35% was shown in L. acidophilus to E. coli, L. acidophilus to C. jejuni was 99.97% to 95.94% and low percentage of L. acidophilus to S. typhi 16S rRNA gene copy numbers after the treatment with CHITPA and CHIAGO. Garlic and onion extract chitosan nanoparticles prepared revealed the presence of selected commensal bacteria and acceptable percentage for caecum L. acidophilus to E. coli and C. jejuni in intestine of Rainbow Rooster Chicken.

| [1] | Torok VA, Ophel-Keller K, Hughes RJ, et al. (2007) Environment and age impact on poultry gut microflora. Aust Poult Sci Symp 19: 149-152. |
| [2] |
Fuller R (1984) Microbial activity in the alimentary tract of birds. Proc Nutr Soc 43: 55-61. doi: 10.1079/PNS19840027

|
| [3] |
Salminen S, Bouley C, Boutron-Ruault MC, et al. (1998) Functional food science and gastrointestinal physiology function. Br J Nutr 80: S147-S171. doi: 10.1079/BJN19980108
|
| [4] |
Hooper LV, Midtvedt T, Gordon JI (2002) How host-microbial interactions shape the nutrient environment of the mammalian intestine. Annu Rev Nutr 22: 283-307. doi: 10.1146/annurev.nutr.22.011602.092259
|
| [5] |
Rehman HU, Vahjen W, Awad WA, et al. (2007) Indigenous bacteria and bacterial metabolic products in the gastrointestinal tract of broiler chickens. Arch Anim Nutr 61: 319-335. doi: 10.1080/17450390701556817
|
| [6] |
Hughes RJ (2008) Relationship between digesta transit time and apparent metabolisable energy value of wheat in chickens. Br Poult Sci 49: 716-720. doi: 10.1080/00071660802449145
|
| [7] |
Pan D, Yu Z (2014) Intestinal microbiome of poultry and its interaction with host and diet. Gut Microbes 5: 108-119. doi: 10.4161/gmic.26945
|
| [8] |
Adil S, Magray SN (2012) Impact and Manipulation of Gut Microflora in Poultry: A Review. J Anim Vet Adv 11: 873-877. doi: 10.3923/javaa.2012.873.877
|
| [9] |
Amit-Romach E, Sklan D, Uni Z (2004) Microflora Ecology of the Chicken Intestine Using 16S Ribosomal DNA Primers. Poult Sci 83: 1093-1098. doi: 10.1093/ps/83.7.1093
|
| [10] |
Vieco-Saiz N, Belguesmia Y, Raspoet R, et al. (2019) Benefits and Inputs from Lactic Acid Bacteria and Their Bacteriocins as Alternatives to Antibiotic Growth Promoters During Food-Animal Production. Front Microbiol 10: 57. doi: 10.3389/fmicb.2019.00057
|
| [11] |
Xu ZR, Hu CH, Xia MS, et al. (2003) Effects of dietary fructooligosaccharide on digestive enzyme activities, intestinal microflora and morphology of male broilers. Poult Sci 82: 1030-1036. doi: 10.1093/ps/82.6.1030
|
| [12] |
Hermans D, Pasmans F, Messens W, et al. (2012) Poultry as a host for the zoonotic pathogen Campylobacter jejuni. Vector-Borne Zoonotic Dis 12: 89-98. doi: 10.1089/vbz.2011.0676
|
| [13] | Albazaz RI, Bal EBB (2004) Microflora of Digestive Tract in Poultry. KSU J Nat Sci 17: 2014. |
| [14] |
Danzeisen JL, Kim HB, Isaacson RE, et al. (2011) Modulations of the Chicken Cecal Microbiome and Metagenome in Response to Anticoccidial and Growth Promoter Treatment. Plos One 6: e27949. doi: 10.1371/journal.pone.0027949
|
| [15] |
Lutful KSM (2009) The role of probiotics in the poultry industry. Int J Mol Sci 10: 3531-3546. doi: 10.3390/ijms10083531
|
| [16] |
Adaszynska MA, Szczerbińska D (2016) Use of essential oils in broiler chicken production – A review. Ann Anim Sci 17: 317-335. doi: 10.1515/aoas-2016-0046
|
| [17] |
Giannenas E, Bonos I, Skoufos A, et al. (2018) Effect of herbal feed additives on performance parameters, intestinal microbiota, intestinal morphology and meat lipid oxidation of broiler chickens. Br Poult Sci 59: 545-553. doi: 10.1080/00071668.2018.1483577
|
| [18] | Rashid Z, Mirani ZA, Zehra S, et al. (2019) Dietary inclusion of phytogenic feed additives (PFA) as alternative to in-feed antibiotics modulating gut microbial dynamics in broiler chicken. Biologia (Pakistan) 65. |
| [19] |
Yadav S, Jha R (2019) Strategies to modulate the intestinal microbiota and their effects on nutrient utilization, performance, and health of poultry. J Anim Sci Biotechnol 10: 2. doi: 10.1186/s40104-018-0310-9
|
| [20] |
Gangadoo S, Stanley D, Hughes RJ, et al. (2016) Nanoparticles in feed: Progress and prospects in poultry research. Trends Food Sci Technol 58: 115-126. doi: 10.1016/j.tifs.2016.10.013
|
| [21] | Huzaifa U, Labaran I, Bello AB, et al. (2014) Phytochemical Screening of Aqueous Extract of Garlic (Allium sativum) bulbs. Rep Opin 6: 1-4. |
| [22] | Viswanathan V, Phadatare AG, Mukne A (2014) Antimycobacterial and Antibacterial Activity of Allium sativum Bulbs. Indian J Pharm Sci 76: 256-261. |
| [23] | Oyebode JA, Fajilade TO (2014) Antibacterial Activities of Aqueous and Ethanolic Extract of Allium Cepa (Onion Bulb) Against Some Selected Pathogenic Microorganisms. Int J Sci Res Publ 4: 2250-3153. |
| [24] |
Mujic I, Dudaš S, Zivkovic J, et al. (2009) Determination of total phenolic compounds in cultivars of Castanea Sativa Mill. Acta Hortic 815: 63-68. doi: 10.17660/ActaHortic.2009.815.8
|
| [25] | Rasaee I, Ghannadnia M, Honari H (2016) Antibacterial properties of biologically formed chitosan nanoparticles using aqueous leaf extract of Ocimum basilicum. Nanomed J 3: 147-154. |
| [26] |
Laudadio V, Passantino L, Perillo A, et al. (2012) Productive performance and histological features of intestinal mucosa of broiler chickens fed different dietary protein levels. Poult Sci 91: 265-270. doi: 10.3382/ps.2011-01675
|
| [27] |
Untergasser A, Nijveen H, Rao X, et al. (2007) Primer3Plus, an enhanced web interface to Primer3. Nucleic Acids Res 35: W71-W74. doi: 10.1093/nar/gkm306
|
| [28] |
Kalendar R, Lee D, Schulman A (2014) FastPCR software for PCR, in silico PCR, and oligonucleotide assembly and analysis. Methods Mol Biol 1116: 271-302. doi: 10.1007/978-1-62703-764-8_18
|
| [29] |
Zhu XY, Zhong T, Pandya Y, et al. (2002) 16S rRNA-based analysis of microbiota from the cecum of broiler chickens. Appl Environ Microbiol 68: 124-137. doi: 10.1128/AEM.68.1.124-137.2002
|
| [30] |
Kralik P, Ricchi M (2017) A Basic Guide to Real Time PCR in Microbial Diagnostics: Definitions, Parameters, and Everything. Front Microbiol 8: 108. doi: 10.3389/fmicb.2017.00108
|
| [31] |
Bustin SA, Benes V, Garson JA, et al. (2009) The MIQE guidelines: minimum information for publication of quantitative real-time PCR experiments. Clin Chem 55: 611-622. doi: 10.1373/clinchem.2008.112797
|
| [32] |
Raymaekers M, Smets R, Maes B, et al. (2009) Checklist for Optimization and Validation of Real-Time PCR Assays. J Clin Lab Anal 23: 145-151. doi: 10.1002/jcla.20307
|
| [33] |
Gotia HT, Munro JB, Knowles DP, et al. (2016) Absolute Quantification of the Host-To-Parasite DNA Ratio in Theileria parva-Infected Lymphocyte Cell Lines. Plos One 11: e0150401. doi: 10.1371/journal.pone.0150401
|
| [34] | Dwight Z, Wittwer CInterpreting melt curves: An indicator, not a diagnosis. (2016) . |
| [35] |
Rychlik W (1995) Selection of primers for polymerase chain reaction. Mol Biotechnol 3: 129-134. doi: 10.1007/BF02789108
|
| [36] |
Ballou AL, Ali RA, Mendoza MA, et al. (2016) Development of the Chick Microbiome: How Early Exposure Influences Future Microbial Diversity. Front Vet Sci 3: 2. doi: 10.3389/fvets.2016.00002
|
| [37] |
Luu QH, Tran TH, Phung DC, et al. (2006) Study on the prevalence of Campylobacter spp. from chicken meat in Hanoi, Vietnam. Ann N Y Acad Sci 1081: 273-275. doi: 10.1196/annals.1373.036
|
| [38] |
Lu X, Rasco BA, Jabal JM, et al. (2011) Investigating antibacterial effects of garlic (Allium sativum) concentrate and garlic-derived organosulfur compounds on Campylobacter jejuni by using Fourier transform infrared spectroscopy, Raman spectroscopy, and electron microscopy. Appl Environ Microbiol 77: 5257-5269. doi: 10.1128/AEM.02845-10
|
| [39] |
Robyn J, Rasschaert G, Hermans D, et al. (2013) Is allicin able to reduce Campylobacter jejuni colonization in broilers when added to drinking water? Poult Sci 92: 1408-1418. doi: 10.3382/ps.2012-02863
|
| [40] |
Kothari D, Lee WD, Niu KM, et al. (2019) The Genus Allium as Poultry Feed Additive: A Review. Animals 9: 1032. doi: 10.3390/ani9121032
|
| [41] |
Curtello S, Vaillant AJ, Asemota H (2018) The Use of Indigenous Plants in the Attenuation of a Live-Attenuated Salmonella Vaccine to Protect Against Poultry. Immunome Res 14: 150. doi: 10.4172/1745-7580.1000150
|
| [42] |
Menconi A, Pumford NR, Morgan MJ, et al. (2014) Effect of chitosan on Salmonella Typhimurium in broiler chickens. Foodborne Pathog Dis 11: 165-169. doi: 10.1089/fpd.2013.1628
|
| [43] | Meinersmann RJ, Rigsby WE, Stern NJ, et al. (1991) Comparative study of colonizing and noncolonizing Campylobacter jejuni. Am J Vet Res 52: 1518-1522. |
| [44] |
Brankatschk R, Bodenhausen N, Zeyer J, et al. (2012) Simple absolute quantification method correcting for quantitative PCR efficiency variations for microbial community samples. Appl Environ Microbiol 78: 4481-4489. doi: 10.1128/AEM.07878-11
|
Figures(7) / Tables(2)

Viviana Imbua Levi Enoka, Gideon Mutie Kikuvi, Perpetual Wangui Ndung'u. Effect of garlic and onion extract chitosan nanoparticles on selected intestinal bacterial flora in indigenous rainbow rooster chicken in Kenya[J]. AIMS Molecular Science, 2021, 8(2): 98-116. doi: 10.3934/molsci.2021008







 DownLoad:
DownLoad: