Citation: AN Mahamad Irfanulla Khan, CS Prashanth, N Srinath. Genetic etiology of cleft lip and cleft palate[J]. AIMS Molecular Science, 2020, 7(4): 328-348. doi: 10.3934/molsci.2020016

| [1] |
Schutte BC, Murray JC (1999) The many faces and factors of orofacial clefts. Hum Mol Genet 8: 1853-1859. doi: 10.1093/hmg/8.10.1853

|
| [2] |
Bender PL (2000) Genetics of cleft lip and palate. J Pediatr Nurs 15: 242-249. doi: 10.1053/jpdn.2000.8148
|
| [3] |
Spritz RA (2001) The genetics and epigenetics of Orofacial clefts. Curr Opin Pediatr 13: 556-560. doi: 10.1097/00008480-200112000-00011
|
| [4] |
Leslie EJ, Marazita ML (2013) Genetics of Cleft Lip and Cleft Palate. Am J Med Genet C Semin Med Genet 163: 246-258. doi: 10.1002/ajmg.c.31381
|
| [5] |
Stanier P, Moore GE (2004) Genetics of cleft lip and palate: syndromic genes contribute to the incidence of non-syndromic clefts. Hum Mol Genet 13: R73-81. doi: 10.1093/hmg/ddh052
|
| [6] |
Mossey P, Little J (2009) Addressing the challenges of cleft lip and palate research in India. Ind J Plast Surg (supplement 1) 42: S9-S18. doi: 10.4103/0970-0358.57182
|
| [7] |
Dixon MJ, Marazita ML, Beaty TH, et al. (2011) Cleft lip and palate: understanding genetic and environmental influences. Nat Rev Genet 12: 167-178. doi: 10.1038/nrg2933
|
| [8] |
Mossey PA, Little J, Munger RG, et al. (2009) Cleft lip and palate. Lancet 374: 1773-1785. doi: 10.1016/S0140-6736(09)60695-4
|
| [9] |
Croen LA, Shaw GM, Wasserman CR, et al. (1998) Racial and ethnic variations in the prevalence of orofacial clefts in California, 1983–1992. Am J Med Genet 79: 42-47. doi: 10.1002/(SICI)1096-8628(19980827)79:1<42::AID-AJMG11>3.0.CO;2-M
|
| [10] | Ching GHS, Chung CS (1974) A genetic study of cleft lip and palate in Hawaii. 1. Interracial crosses. Am J Hum Genet 26: 162-172. |
| [11] | Murray JC, Daack-Hirsch S, Buetow KH, et al. (1997) Clinical and epidemiologic studies of cleft lip and palate in the Philippines. Cleft Palate Craniofac J 34: 7-10. |
| [12] | Vanderas AP (1987) Incidence of cleft lip, cleft palate, and cleft lip and palate among races: a review. Cleft Palate J 24: 216-225. |
| [13] |
Hagberg C, Larson O, Milerad J (1997) Incidence of cleft lip and palate and risks of additional malformations. Cleft Palate Craniofac J 35: 40-45. doi: 10.1597/1545-1569_1998_035_0040_ioclap_2.3.co_2
|
| [14] | Ogle OE (1993) Incidence of cleft lip and palate in a newborn Zairian sample. Cleft Palate Craniofac J 30: 250-251. |
| [15] |
Chung CS, Mi MP, Beechert AM, et al. (1987) Genetic epidemiology of cleft lip with or without cleft palate in the population of Hawaii. Genetic Epidemiol 4: 415-423. doi: 10.1002/gepi.1370040603
|
| [16] |
Wyszynski DF, Wu T (2002) Prenatal and perinatal factors associated with isolated oral clefting. Cleft Palate Craniofac J 39: 370-375. doi: 10.1597/1545-1569_2002_039_0370_papfaw_2.0.co_2
|
| [17] |
Mossey PA (2007) Epidemiology underpinning research in the etiology of orofacial clefts. Orthod Craniofac Res 10: 114-120. doi: 10.1111/j.1601-6343.2007.00398.x
|
| [18] |
Yang J, Carmichael SL, Canfield M, et al. (2008) Socioeconomic status in relation to selected birth defects in a large multicentered US case-control study. Am J Epidemiol 167: 145-154. doi: 10.1093/aje/kwm283
|
| [19] |
Sabbagh HJ, Innes NP, Sallout BI, et al. (2015) Birth prevalence of non-syndromicorofacial clefts in Saudi Arabia and the effects of parental consanguinity. Saudi Med J 36: 1076-1083. doi: 10.15537/smj.2015.9.11823
|
| [20] |
Silva CM, Pereira MCM, Queiroz TB, et al. (2019) Can Parental Consanguinity Be a Risk Factor for the Occurrence of Nonsyndromic Oral Cleft? Early Hum Dev 135: 23-26. doi: 10.1016/j.earlhumdev.2019.06.005
|
| [21] |
Sabbagh HJ, Hassan MH, Innes NP, et al. (2014) Parental Consanguinity and Nonsyndromic Orofacial Clefts in Children: A Systematic Review and Meta-Analyses. Cleft Palate Craniofac J 51: 501-513. doi: 10.1597/12-209
|
| [22] |
Reddy SG, Reddy RR, Bronkhorst EM, et al. (2010) Incidence of cleft lip and palate in the state of Andhra Pradesh, South India. Indian J Plast Surg 43: 184-189. doi: 10.4103/0970-0358.73443
|
| [23] | Indian Council of Medical Research (ICMR) Task Force Project (2016) Division of Non-Communicable Diseases; New Delhi. |
| [24] |
Neela PK, Reddy SG, Husain A, et al. (2019) Association of cleft lip and/or palate in people born to consanguineous parents: A 13year retrospective study from a very high volume cleft center. J Cleft Lip Palate Craniofac Anoma l6: 33-37. doi: 10.4103/jclpca.jclpca_34_18
|
| [25] |
Babu GV, Syed AH, Murthy J, et al. (2018) IRF6 rs2235375 single nucleotide polymorphism is associated with isolated non-syndromic cleft palate but not with cleft lip with or without palate in South Indian population. Braz J Otorhinolaryngol 84: 473-477. doi: 10.1016/j.bjorl.2017.05.011
|
| [26] |
Babu GV, Syed AH, Murthy J, et al. (2015) Evidence of the involvement of the polymorphisms near MSX1 gene in non-syndromic cleft lip with or without cleft palate. Int J Pediatr Otorhinolaryngol 79: 1081-1084. doi: 10.1016/j.ijporl.2015.04.034
|
| [27] |
Neela PK, Reddy SG, Husain A, et al. (2020) CRISPLD2 Gene Polymorphisms with Nonsyndromic Cleft Lip and Palate in Indian Population. Global Med Genet 7: 22-25. doi: 10.1055/s-0040-1713166
|
| [28] |
Tolarova MM, Cervenka J (1998) Classification and birth prevalence of Orofacial clefts. Am J Med Genet 75: 126-137. doi: 10.1002/(SICI)1096-8628(19980113)75:2<126::AID-AJMG2>3.0.CO;2-R
|
| [29] | Wong FK, Hagg U (2004) An update on the aetiology of orofacial clefts. Hong Kong Med J 10: 331-336. |
| [30] |
Rahimov F, Jugessur A, Murray JC (2012) Genetics of Nonsyndromic Orofacial Clefts. Cleft Palate Craniofac J 49: 73-91. doi: 10.1597/10-178
|
| [31] | Jones MC (1988) Etiology of facial clefts. Prospective evaluation of 428 patients. Cleft Palate J 25: 16-20. |
| [32] | Kohli SS, Kohli VS (2012) A comprehensive review of the genetic basis of cleft lip and palate. J Oral MaxillofacPathol 16: 64-72. |
| [33] |
Wyszynski DF, Wu T (2002) Prenatal and perinatal factors associated with isolated oral clefting. Cleft Palate Craniofac J 39: 370-375. doi: 10.1597/1545-1569_2002_039_0370_papfaw_2.0.co_2
|
| [34] | Cobourne MT (2012) Cleft Lip and Palate. Epidemiology, Aetiology and Treatment. Front Oral Biol London: Karger, 60-70. |
| [35] | Gundlach KK, Maus C (2006) Epidemiological studies on the frequency of clefts in Europe and world-wide. J Craniomaxillofac Surg 34: 1-2. |
| [36] | Hobbs J (1991) The adult patient. Clin Commun Disord 1: 48-52. |
| [37] |
Murthy J (2009) Management of cleft lip and palate in adults. Indian J Plast Surg 42: S116-S122. doi: 10.4103/0970-0358.57202
|
| [38] |
Murray JC (2002) Gene/environment causes of cleft lip and/or palate. Clin Genet 61: 248-256. doi: 10.1034/j.1399-0004.2002.610402.x
|
| [39] |
Funato N, Nakamura M (2017) Identification of shared and unique gene families associated with oral clefts. Int J Oral Sci 9: 104-109. doi: 10.1038/ijos.2016.56
|
| [40] |
Lie RT, Wilcox AJ, Taylor J, et al. (2008) Maternal smoking and oral clefts: the role of detoxification pathway genes. Epidemiology 19: 606-615. doi: 10.1097/EDE.0b013e3181690731
|
| [41] |
Stothard KJ, Tennant PW, Bell R, et al. (2009) Maternal overweight and obesity and the risk of congenital anomalies: a systematic review and meta-analysis. JAMA 301: 636-650. doi: 10.1001/jama.2009.113
|
| [42] |
Van Rooij IA, Ocké MC, Straatman H, et al. (2004) Periconceptional folate intake by supplement and food reduces the risk of nonsyndromic cleft lip with or without cleft palate. Prev Med 39: 689-694. doi: 10.1016/j.ypmed.2004.02.036
|
| [43] |
Wilcox AJ, Lie RT, Solvoll K, et al. (2007) Folic acid supplements and risk of facial clefts: national population based case-control study. BMJ 334: 464. doi: 10.1136/bmj.39079.618287.0B
|
| [44] |
Martyn TC (2004) The complex genetics of cleft lip and palate. Eur J Orthod 26: 7-16. doi: 10.1093/ejo/26.1.7
|
| [45] | Hozyasz KK, Mostowska A, Surowiec Z, et al. (2005) Genetic polymorphisms of GSTM1 and GSTT1 in mothers of children with isolated cleft lip with or without cleft palate. Przegl Lek 62: 1019-1022. |
| [46] |
Junaid M, Narayanan MBA, Jayanthi D, et al. (2018) Association between maternal exposure to tobacco, presence of TGFA gene, and the occurrence of oral clefts. A case control study. Clin Oral Investig 22: 217-223. doi: 10.1007/s00784-017-2102-6
|
| [47] |
Chevrier C, Perret C, Bahuau M, et al. (2005) Interaction between the ADH1C polymorphism and maternal alcohol intake in the risk of nonsyndromic oral clefts: an evaluation of the contribution of child and maternal genotypes. Birth Defects Res A Clin Mol Teratol 73: 114-122. doi: 10.1002/bdra.20103
|
| [48] |
Jugessur A, Shi M, Gjessing HK, et al. (2009) Genetic determinants of facial clefting: analysis of 357 candidate genes using two national cleft studies from Scandinavia. PLoS One 4: e5385. doi: 10.1371/journal.pone.0005385
|
| [49] | Mossey PA, Little J (2002) Epidemiology of oral clefts: An international perspective. Cleft lip and palate: From origin to treatment Oxford: Oxford University Press, 127-144. |
| [50] |
Krapels IP, van Rooij IA, Ocke MC, et al. (2004) Maternal nutritional status and the risk for orofacial cleft offspring in humans. J Nutr 134: 3106-3113. doi: 10.1093/jn/134.11.3106
|
| [51] |
Hayes C, Werler MM, Willett WC, et al. (1996) Case-control study of periconceptional folic acid supplementation and orofacial clefts. Am J Epidemiol 143: 1229-1234. doi: 10.1093/oxfordjournals.aje.a008710
|
| [52] |
Thomas D (2010) Gene-environment-wide association studies: emerging approaches. Nat Rev Genet 11: 259-272. doi: 10.1038/nrg2764
|
| [53] |
Zhao M, Ren Y, Shen L, et al. (2014) Association between MTHFR C677T and A1298C Polymorphisms and NSCL/P Risk in Asians: A Meta-Analysis. PLoS One 9: e88242. doi: 10.1371/journal.pone.0088242
|
| [54] |
Jagomägi T, Nikopensius T, Krjutškov K, et al. (2010) MTHFR and MSX1 contribute to the risk of nonsyndromic cleft lip/palate. Eur J Oral Sci 118: 213-220. doi: 10.1111/j.1600-0722.2010.00729.x
|
| [55] |
Borkar AS (1993) Epidemiology of facial clefts in the central province of Saudi Arabia. Br J Plast Surg 46: 673-675. doi: 10.1016/0007-1226(93)90198-K
|
| [56] | Burdick AB (1986) Genetic epidemiology and control of genetic expression in Van der Woude syndrome. J Craniofac Genet Dev Biol Suppl 2: 99-105. |
| [57] |
Kondo S, Schutte BC, Richardson RJ, et al. (2002) Mutations in IRF6 cause Van der Woude and popliteal pterygium syndromes. Nat Genet 32: 285-289. doi: 10.1038/ng985
|
| [58] |
Zucchero TM, Cooper ME, Maher BS, et al. (2004) Interferon regulatory factor 6 (IRF6) gene variantsand the risk of isolated cleft lip or palate. N Engl J Med 351: 769-780. doi: 10.1056/NEJMoa032909
|
| [59] |
Blanton SH, Cortez A, Stal S, et al. (2005) Variation in IRF6 contributes to nonsyndromic cleft lip and palate. Am J Med Genet A 137: 259-262. doi: 10.1002/ajmg.a.30887
|
| [60] |
Jugessur A, Rahimov F, Lie RT, et al. (2008) Genetic variants in IRF6 and the risk of facial clefts: single-marker and haplotype-based analyses in a population-based case-control study of facial clefts in Norway. Genet Epidemiol 32: 413-424. doi: 10.1002/gepi.20314
|
| [61] |
Huang Y, Wu J, Ma J, et al. (2009) Association between IRF6 SNPs and oral clefts in West China. J Dent Res 88: 715-718. doi: 10.1177/0022034509341040
|
| [62] |
Grant SF, Wang K, Zhang H, et al. (2009) A Genome-Wide Association Study Identifies a Locus for Nonsyndromic Cleft Lip with or without Cleft Palate on 8q24. J Pediatr 155: 909-913. doi: 10.1016/j.jpeds.2009.06.020
|
| [63] |
Beaty TH, Murray JC, Marazita ML, et al. (2010) A genome-wide association study of cleft lip with and without cleft palate identifies risk variants near MAFB and ABCA4. Nat Genet 42: 525-529. doi: 10.1038/ng.580
|
| [64] |
Ibarra-Arce A, García-Álvarez M, Cortés-González D (2015) IRF6 polymorphisms in Mexican patients with non-syndromic cleft lip. Meta Gene 4: 8-16. doi: 10.1016/j.mgene.2015.02.002
|
| [65] |
Satokata I, Maas R (1994) Msx1 deficient mice exhibit cleft palate and abnormalities of craniofacial and tooth development. Nat Genet 6: 348-356. doi: 10.1038/ng0494-348
|
| [66] |
Van den Boogaard MJ, Dorland M, Beemer FA, et al. (2000) MSX1 mutation is associated with Orofacial clefting and tooth agenesis in humans. Nat Genet 24: 342-343. doi: 10.1038/74155
|
| [67] |
Jezewski PA, Vieira AR, Nishimura C, et al. (2003) Complete sequencing shows a role for MSX1 in nonsyndromic cleft lip and palate. J Med Genet 40: 399-407. doi: 10.1136/jmg.40.6.399
|
| [68] |
Otero L, Gutierrez S, Chaves M, et al. (2007) Association of MSX1 with nonsyndromic cleft lip and palate in a Colombian population. Cleft palate craniofac J 44: 653-656. doi: 10.1597/06-097.1
|
| [69] |
Salahshourifar I, Halim AS, Sulaiman WA, et al. (2011) Contribution of MSX1 variants to the risk of non-syndromic cleft lip and palate in a Malay population. J Hum Genet 56: 755-758. doi: 10.1038/jhg.2011.95
|
| [70] | Indencleef K, Roosenboom J, Hoskens H, et al. (2018) Six NSCL/P loci show associations with normal-range craniofacial variation. Front. Genet 9: 502. |
| [71] |
Tasanarong P, Pabalan N, Tharabenjasin P, et al. (2019) MSX1 gene polymorphisms and non-syndromic cleft lip with or without palate (NSCL/P): A meta-analysis. Oral Dis 25: 1492-1501. doi: 10.1111/odi.13127
|
| [72] |
Mitchell LE (1997) Transforming growth factor alpha locus and nonsyndromic cleft lip with or without cleft palate: a reappraisal. Genet Epidemiol 14: 231-240. doi: 10.1002/(SICI)1098-2272(1997)14:3<231::AID-GEPI2>3.0.CO;2-8
|
| [73] |
Shaw GM, Wasserman CR, Murray JC, et al. (1998) Infant TGF-alpha genotype, orofacial clefts, and maternal periconceptional multivitamin use. Cleft Palate Craniofac J 35: 366-367. doi: 10.1597/1545-1569_1998_035_0366_itagoc_2.3.co_2
|
| [74] |
Holder SE, Vintiner GM, Farren B, et al. (1999) Confirmation of an association between RFLPs at the transforming growth factor-alpha locus and nonsyndromic cleft lip and palate. J Med Genet 29: 390-392. doi: 10.1136/jmg.29.6.390
|
| [75] |
Passos-Bueno MR, Gaspar DA, Kamiya T, et al. (2004) Transforming growth factor-alpha and non-syndromic cleft lip with or without palate in Brazilian patients: results of large case-control study. Cleft Palate Craniofac J 41: 387-391. doi: 10.1597/03-054.1
|
| [76] |
Letra A, Fakhouri W, Renata F, et al. (2012) Interaction between IRF6 and TGFA Genes Contribute to the Risk of Nonsyndromic Cleft Lip/Palate. PLoS One 7: e45441. doi: 10.1371/journal.pone.0045441
|
| [77] | Feng C, Zhang E, Duan W, et al. (2014) Association Between Polymorphism of TGFA Taq I and Cleft Lip and/or Palate: A Meta-Analysis. BMC Oral Health 11: 14-88. |
| [78] |
Yan C, He DQ, Chen LY, et al. (2018) Transforming Growth Factor Alpha Taq I Polymorphisms and Nonsyndromic Cleft Lip and/or Palate Risk: A Meta-Analysis. Cleft Palate Craniofac J 55: 814-820. doi: 10.1597/16-008
|
| [79] |
Suzuki S, Marazita ML, Cooper ME, et al. (2009) Mutations in BMP4 are associated with subepithelial, microform, and overt cleft lip. Am J Hum Genet 84: 406-411. doi: 10.1016/j.ajhg.2009.02.002
|
| [80] |
Antunes LS, Küchler EC, Tannure PN, et al. (2013) BMP4 Polymorphism is Associated with Nonsyndromic Oral Cleft in a Brazilian Population. Cleft Palate Craniofac J 50: 633-638. doi: 10.1597/12-048
|
| [81] |
Li YH, Yang J, Zhang JL, et al. (2017) BMP4 rs17563 polymorphism and nonsyndromic cleft lip with or without cleft palate: a meta- analysis. Medicine (Baltim) 96: e7676. doi: 10.1097/MD.0000000000007676
|
| [82] |
Hao J, Gao R, Wua W, et al. (2018) Association between BMP4 gene polymorphisms and cleft lip with or without cleft palate in a population from South China. Arch Oral Biol 93: 95-99. doi: 10.1016/j.archoralbio.2018.05.015
|
| [83] |
Assis Machado R, De Toledo IP, Martelli-Junior H, et al. (2018) Potential genetic markers for nonsyndromic oral clefts in the Brazilian population: a systematic review and meta-analysis. Birth Defects Res 110: 827-839. doi: 10.1002/bdr2.1208
|
| [84] | Mansouri A, Stoykova A, Torres M, et al. (1996) Dysgenesis of cephalic neural crest derivatives in Pax7 mutant mice. Development 122: 831-838. |
| [85] |
Sull JW, Liang KY, Hetmanski JB, et al. (2009) Maternal transmission effects of the PAX genes among cleft case-parent trios from four populations. Eur J Hum Genet 17: 831-839. doi: 10.1038/ejhg.2008.250
|
| [86] |
Ludwig KU, Mangold E, Herms S, et al. (2012) Genome-wide meta-analyses of non syndromic cleft lip with or without cleft palate identify six new risk loci. Nat Genet 44: 968-971. doi: 10.1038/ng.2360
|
| [87] |
Beaty TH, Taub MA, Scott AF, et al. (2013) confirming genes influencing risk to Cleft lip with or without cleft palate in a case – parent trio study. Hum Genet 132: 771-781. doi: 10.1007/s00439-013-1283-6
|
| [88] |
Leslie EJ, Taub MA, Liu H, et al. (2015) Identification of Functional Variants for Cleft Lip with or without Cleft Palate in or near PAX7, FGFR2 and NOG by Targeted Sequencing of GWAS Loci. Am J Hum Genet 96: 397-411. doi: 10.1016/j.ajhg.2015.01.004
|
| [89] | Duan SJ, Huang N, Zhang BH, et al. (2017) New insights from GWAS for the cleft palate among han Chinese population. Med Oral Patol Oral Cir Bucal 22: e219-e227. |
| [90] |
Gaczkowska A, Biedziak B, Budner M, et al. (2019) PAX7 nucleotide variants and the risk of non-syndromicorofacial clefts in the Polish population. Oral Dis 25: 1608-1618. doi: 10.1111/odi.13139
|
| [91] |
Otto F, Kanegane H, Mundlos S (2002) Mutations in the RUNX2 gene in patients with cleidocranial dysplasia. Hum Mutat 19: 209-216. doi: 10.1002/humu.10043
|
| [92] |
Cooper SC, Flaitz CM, Johnston DA, et al. (2001) A natural history of cleidocranial dysplasia. Am J Med Genet 104: 1-6. doi: 10.1002/ajmg.10024
|
| [93] |
Aberg T, Cavender A, Gaikwad JS, et al. (2004) Phenotypic changes in dentition of Runx2 homozygote-null mutant mice. J Histochem Cytochem 52: 131. doi: 10.1177/002215540405200113
|
| [94] |
Sull JW, Liang KY, Hetmanski JB, et al. (2008) Differential parental transmission of markers in RUNX2 among cleft case-parent trios from four populations. Genet Epidemiol 32: 505-512. doi: 10.1002/gepi.20323
|
| [95] |
Wu T, Daniele M, Fallin MD, et al. (2012) Evidence of Gene-Environment Interaction for the RUNX2 Gene and Environmental Tobacco Smoke in Controlling the Risk of Cleft Lip with/without Cleft Palate. Birth Defects Res A Clin Mol Teratol 94: 76-83. doi: 10.1002/bdra.22885
|
| [96] |
Alkuraya FS, Saadi I, Lund JJ, et al. (2006) SUMO1 haploinsufficiency leads to cleft lip and palate. Science 313: 1751. doi: 10.1126/science.1128406
|
| [97] |
Song T, Li G, Jing G, et al. (2008) SUMO1 polymorphisms are associated with non-syndromic cleft lip with or without cleft palate. Biochem Biophys Res Commun 377: 1265-1268. doi: 10.1016/j.bbrc.2008.10.138
|
| [98] | Carter TC, Molloy AM, Pangilinan F, et al. (2010) Testing reported associations of genetic risk factors for oral clefts in a large Irish study population. Birth Defects Res A Clin Mol Teratol 88: 84-93. |
| [99] |
Tang MR, Wang YX, Han SY, et al. (2014) SUMO1 genetic polymorphisms may contribute to the risk of nonsyndromic cleft lip with or without palate: a meta-analysis. Genet Test Mol Biomarkers 18: 616-624. doi: 10.1089/gtmb.2014.0056
|
| [100] |
Hallonet M, Hollemann T, Pieler T, et al. (1999) VAX1, a novel homeobox-containing gene, directs development of the basal forebrain and visual system. Genes Dev 13: 3106-3114. doi: 10.1101/gad.13.23.3106
|
| [101] |
Slavotinek AM, Chao R, Vacik T, et al. (2012) VAX1 mutation associated with microphthalmia, corpus callosum agenesis, and orofacial clefting: the first description of a VAX1 phenotype in humans. Hum Mutat 33: 364-368. doi: 10.1002/humu.21658
|
| [102] |
Butali A, Suzuki S, Cooper ME, et al. (2013) Replication of Genome Wide Association Identified Candidate Genes Confirm the Role of Common and Rare Variants in PAX7 and VAX1 in the Etiology of Non-syndromic CL/P. Am J Med Genet A 161: 965-972. doi: 10.1002/ajmg.a.35749
|
| [103] |
Zhang BH, Shi JY, Lin YS, et al. (2018) VAX1 Gene Associated Non-Syndromic Cleft Lip With or Without Palate in Western Han Chinese. Arch Oral Biol 95: 40-43. doi: 10.1016/j.archoralbio.2018.07.014
|
| [104] |
Sabbagh HJ, Innes NPT, Ahmed ES, et al. (2019) Molecular Screening of VAX1 Gene Polymorphisms Uncovered the Genetic Heterogeneity of Nonsyndromic Orofacial Cleft Among Saudi Arabian Patients. Genet Test Mol Biomarkers 23: 45-50. doi: 10.1089/gtmb.2018.0207
|
| [105] |
De Felice M, Ovitt C, Biffali E, et al. (1998) A mouse model for hereditary thyroid dysgenesis and cleft palate. Nature Genet 19: 395-398. doi: 10.1038/1289
|
| [106] |
Bamforth JS, Hughes IA, Lazarus JH, et al. (1989) Congenital hypothyroidism, spiky hair, and cleft palate. J Med Genet 26: 49-60. doi: 10.1136/jmg.26.1.49
|
| [107] |
Castanet M, Park SM, Smith A, et al. (2002) A novel loss-of-function mutation in TTF-2 is associated with congenital hypothyroidism, thyroid agenesis and cleft palate. Hum Mol Genet 11: 2051-2059. doi: 10.1093/hmg/11.17.2051
|
| [108] |
Nikopensius T, Kempa I, Ambrozaitytė L, et al. (2011) Variation in FGF1, FOXE1, and TIMP2 genes is associated with nonsyndromic cleft lip with or without cleft palate. Birth Defects Re Part A 91: 218-225. doi: 10.1002/bdra.20791
|
| [109] |
Leslie EJ, Carlson JC, Shaffer JR, et al. (2017) Genome-wide meta-analyses of nonsyndromicorofacial clefts identify novel associations between FOXE1 and all orofacial clefts, and TP63 and cleft lip with or without cleft palate. Hum Genet 136: 275-286. doi: 10.1007/s00439-016-1754-7
|
| [110] |
Chiquet BT, Lidral AC, Stal S, et al. (2007) CRISPLD2: a novel NSCLP candidate gene. Hum Mol Genet 16: 2241-2248. doi: 10.1093/hmg/ddm176
|
| [111] |
Shi J, Jiao X, Song T, et al. (2010) CRISPLD2 polymorphisms are associated with nonsyndromic cleft lip with or without cleft palate in a northern Chinese population. Eur J Oral Sci 118: 430-433. doi: 10.1111/j.1600-0722.2010.00743.x
|
| [112] |
Letra A, Menezes R, Margaret E, et al. (2011) CRISPLD2 Variants Including a C471T Silent Mutation May Contribute to Nonsyndromic Cleft Lip with or without Cleft Palate. Cleft Palate Craniofac J 48: 363-370. doi: 10.1597/09-227
|
| [113] |
Mijiti A, Ling W, Maimaiti A, et al. (2015) Preliminary evidence of an interaction between the CRISPLD2 gene and non-syndromic cleft lip with or without cleft palate (nsCL/P) in Xinjiang Uyghur population, China. Int J Pediatr Otorhinolaryngol 79: 94-100. doi: 10.1016/j.ijporl.2014.10.043
|
| [114] |
Chiquet BT, Yuan Q, Swindell EC, et al. (2018) Knockdown of Crispld2 in zebrafish identifies a novel network for nonsyndromic cleft lip with or without cleft palate candidate genes. Eur J Hum Genet 26: 1441-1450. doi: 10.1038/s41431-018-0192-5
|
| [115] |
Greene ND, Dunlevy LP, Copp AJ (2003) Homocysteine Is Embryo toxic but Does Not Cause Neural Tube Defects in Mouse Embryos. Anat Embryo 206: 185-191. doi: 10.1007/s00429-002-0284-3
|
| [116] |
Gaspar DA, Matioli SR, Pavanello RC, et al. (2004) Maternal MTHFR Interacts With the Offspring's BCL3 Genotypes, but Not With TGFA, in Increasing Risk to Nonsyndromic Cleft Lip With or Without Cleft Palate. Eur J Hum Genet 12: 521-526. doi: 10.1038/sj.ejhg.5201187
|
| [117] |
Rai V (2018) Strong Association of C677T Polymorphism of Methylenetetrahydrofolate Reductase Gene With Nosyndromic Cleft Lip/Palate (nsCL/P). Indian J Clin Biochem 33: 5-15. doi: 10.1007/s12291-017-0673-2
|
| [118] |
Bhaskar L, Murthy J, Babu GV (2011) Polymorphisms in Genes Involved in Folate Metabolism and Orofacial Clefts. Arch Oral Biol 56: 723-737. doi: 10.1016/j.archoralbio.2011.01.007
|
| [119] |
Machado AR, De Toledo IP, Martelli H, et al. (2018) Potential genetic markers for nonsyndromic oral clefts in the Brazilian population: a systematic review and meta-analysis. Birth Defects Res 110: 827-839. doi: 10.1002/bdr2.1208
|
| [120] |
Lin L, Bu H, Yang Y, et al. (2017) A Targeted, Next-Generation Genetic Sequencing Study on Tetralogy of Fallot, Combined With Cleft Lip and Palate. J Craniofac Surg 28: e351-e355. doi: 10.1097/SCS.0000000000003598
|
| [121] |
Lan Y, Ryan RC, Zhang Z, et al. (2006) Expression of Wnt9b and activation of canonical Wnt signaling during midfacial morphogenesis in mice. Dev Dyn 235: 1448-1454. doi: 10.1002/dvdy.20723
|
| [122] |
Carroll TJ, Park JS, Hayashi S, et al. (2005) Wnt9b plays a central role in the regulation of mesenchymal to epithelial transitions underlying organogenesis of the mammalian urogenital system. Dev Cell 9: 283-292. doi: 10.1016/j.devcel.2005.05.016
|
| [123] |
Karner CM, Chirumamilla R, Aoki S, et al. (2009) Wnt9b signaling regulates planar cell polarity and kidney tubule morphogenesis. Nature Genet 41: 793-799. doi: 10.1038/ng.400
|
| [124] |
Juriloff DM, Harris MJ, McMahon AP, et al. (2006) Wnt9b is the mutated gene involved in multifactorial nonsyndromic cleft lip with or without cleft palate in A/WySn mice, as confirmed by a genetic complementation test. Birth Defects Res A Clin Mol Teratol 76: 574-579. doi: 10.1002/bdra.20302
|
| [125] |
Chiquet BT, Blanton SH, Burt A, et al. (2008) Variation in WNT genes is associated with non-syndromic cleft lip with or without cleft palate. Hum Mol Genet 17: 2212-2218. doi: 10.1093/hmg/ddn121
|
| [126] |
Menezes R, Letra A, Kim AH, et al. (2010) Studies with Wnt genes and nonsyndromic cleft lip and palate. Birth Defects Res A Clin Mol Teratol 88: 995-1000. doi: 10.1002/bdra.20720
|
| [127] |
Fontoura C, Silva RM, Granjeiro JM, et al. (2015) Association of WNT9B Gene Polymorphisms With Nonsyndromic Cleft Lip With or Without Cleft Palate in Brazilian Nuclear Families. Cleft Palate Craniofac J 52: 44-48. doi: 10.1597/13-146
|
| [128] |
Vijayan V, Ummer R, Weber R, et al. (2018) Association of WNT Pathway Genes With Nonsyndromic Cleft Lip With or Without Cleft Palate. Cleft Palate Craniofac J 55: 335-341. doi: 10.1177/1055665617732782
|
| [129] |
Marini NJ, Asrani K, Yang W, et al. (2019) Accumulation of rare coding variants in genes implicated in risk of human cleft lip with or without cleft palate. Am J Med Genet 179: 1260-1269. doi: 10.1002/ajmg.a.61183
|
| [130] |
Gajera M, Desai N, Suzuki A, et al. (2019) MicroRNA-655-3p and microRNA-497-5p inhibit cell proliferation in cultured human lip cells through the regulation of genes related to human cleft lip. BMC Med Genomics 12: 70. doi: 10.1186/s12920-019-0535-2
|
| [131] |
Kim SJ, Lee S, Park HJ, et al. (2016) Genetic association of MYH genes with hereditary hearing loss in Korea. Gene 591: 177-182. doi: 10.1016/j.gene.2016.07.011
|
| [132] |
Pazik J, Lewandowski Z, Oldak M, et al. (2016) Association of MYH9 rs3752462 and rs5756168 polymorphisms with transplanted kidney artery stenosis. Transplant Proc 48: 1561-1565. doi: 10.1016/j.transproceed.2016.01.085
|
| [133] |
Marigo V, Nigro A, Pecci A, et al. (2004) Correlation between the clinical phenotype of MYH9-related disease and tissue distribution of class II nonmuscle myosin heavy chains. Genomics 83: 1125-1133. doi: 10.1016/j.ygeno.2003.12.012
|
| [134] |
Martinelli M, Di Stazio M, Scapoli L, et al. (2007) Cleft lip with or without cleft palate: implication of the heavy chain of non-muscle myosin IIA. J Med Genet 44: 387-392. doi: 10.1136/jmg.2006.047837
|
| [135] |
Chiquet BT, Hashmi SS, Henry R, et al. (2009) Genomic screening identifies novel linkages and provides further evidence for a role of MYH9 in nonsyndromic cleft lip and palate. Eur J Hum Genet 17: 195-204. doi: 10.1038/ejhg.2008.149
|
| [136] |
Jia ZL, Li Y, Chen CH, et al. (2010) Association among polymorphisms at MYH9, environmental factors, and nonsyndromicorofacial clefts in western China. DNA Cell Biol 29: 25-32. doi: 10.1089/dna.2009.0935
|
| [137] |
Peng HH, Chang NC, Chen KT, et al. (2016) Nonsynonymous variants in MYH9 and ABCA4 are the most frequent risk loci associated with nonsyndromicorofacial cleft in Taiwanese population. BMC Med Genet 17: 59. doi: 10.1186/s12881-016-0322-2
|
| [138] |
Wang Y, Li D, Xu Y, et al. (2018) Functional Effects of SNPs in MYH9 and Risks of Nonsyndromic Orofacial Clefts. J Dent Res 97: 388-394. doi: 10.1177/0022034517743930
|
| [139] |
Araujo TK, Secolin R, Félix TM, et al. (2016) A multicentric association study between 39 genes and nonsyndromic cleft lip and palate in a Brazilian population. J Craniomaxillofac Surg 44: 16-20. doi: 10.1016/j.jcms.2015.07.026
|
| [140] |
Yu Y, Zuo X, He M, et al. (2017) Genome-wide analyses of non-syndromic cleft lip with palate identify 14 novel loci and genetic heterogeneity. Nat Commun 8: 14364. doi: 10.1038/ncomms14364
|
| [141] |
Da Silva HPV, Oliveira GHM, Ururahy MAG, et al. (2018) Application of high-resolution array platform for genome-wide copy number variation analysis in patients with nonsyndromic cleft lip and palate. J Clin Lab Anal 32: e22428. doi: 10.1002/jcla.22428
|
| [142] |
Huang L, Jia Z, Shi Y, et al. (2019) Genetic factors define CPO and CLO subtypes of nonsyndromicorofacial cleft. PLoS Genet 15: e1008357. doi: 10.1371/journal.pgen.1008357
|
| [143] | Gorlin RJ, Cohen MM, Hennekam RCM (2001) Syndromes of the Head and Neck New York: Oxford University Press. |
| [144] |
Grosen D, Chevrier C, Skytthe A, et al. (2010) A cohort study of recurrence patterns among more than 54000 relatives of oral cleft cases in Denmark: support for the multifactorial threshold model of inheritance. J Med Genet 47: 162-168. doi: 10.1136/jmg.2009.069385
|
| [145] |
Lidral AC, Romitti PA, Basart AM, et al. (1998) Association of MSX1 and TGFB3 with nonsyndromicclefting in humans. Am J Hum Genet 63: 557-568. doi: 10.1086/301956
|
| [146] |
Suazo J, Santos JL, Scapoli L, et al. (2010) Association between TGFB3 and nonsyndromic cleft lip with or without cleft palate in a Chilean population. Cleft Palate Craniofac J 47: 513-517. doi: 10.1597/09-015
|
| [147] |
Vieira AR, Avila JR, Daack-Hirsch S, et al. (2005) Medical sequencing of candidate genes for nonsyndromic cleft lip and palate. PLoS Genet 1: e64. doi: 10.1371/journal.pgen.0010064
|
| [148] |
Choi SJ, Marazita ML, Hart PS, et al. (2009) The PDGF-C regulatory region SNP rs28999109 decreases promoter transcriptional activity and is associated with CL/P. Eur J Hum Genet 17: 774-784. doi: 10.1038/ejhg.2008.245
|
| [149] |
François-Fiquet C, Poli-Merol ML, Nguyen P, et al. (2014) Role of angiogenesis-related genes in cleft lip/palate: review of the literature. Int J Pediatr Otorhinolaryngol 78: 1579-1585. doi: 10.1016/j.ijporl.2014.08.001
|
| [150] |
Carlson JC, Taub MA, Feingold E, et al. (2017) Identifying Genetic Sources of Phenotypic Heterogeneity in Orofacial Clefts by Targeted Sequencing. Birth Defects Res 109: 1030-1038. doi: 10.1002/bdr2.23605
|
| [151] |
Figueiredo JC, Stephanie LY, Raimondi H, et al. (2014) Genetic risk factors for orofacial clefts in Central Africans and Southeast Asians. Am J Med Genet A 164A: 2572-2580. doi: 10.1002/ajmg.a.36693
|
| [152] |
Leslie EJ, Taub MA, Liu H, et al. (2015) Identification of Functional Variants for Cleft Lip with or without Cleft Palate in or near PAX7, FGFR2, and NOG by Targeted Sequencing of GWAS Loci. Am J Hum Genet 96: 397-411. doi: 10.1016/j.ajhg.2015.01.004
|
| [153] |
Moreno Uribe LM, Fomina T, Munger RG, et al. (2017) A Population-Based Study of Effects of Genetic Loci on Orofacial Clefts. J Dent Res 96: 1322-1329. doi: 10.1177/0022034517716914
|
| [154] |
Liu H, Leslie EJ, Carlson JC, et al. (2017) Identification of common non-coding variants at 1p22 that are functional for non-syndromic orofacial clefting. Nat Commun 8: 14759. doi: 10.1038/ncomms14759
|
| [155] |
Cox LL, Cox TC, Moreno Uribe LM, et al. (2018) Mutations in the Epithelial Cadherin-p120-Catenin Complex Cause Mendelian Non-Syndromic Cleft Lip with or without Cleft Palate. Am J Hum Genet 102: 1143-1157. doi: 10.1016/j.ajhg.2018.04.009
|
| [156] |
Du S, Yang Y, Yi P, et al. (2019) A Novel CDH1 Mutation Causing Reduced E-Cadherin Dimerization Is Associated with Nonsyndromic Cleft Lip With or Without Cleft Palate. Genet Test Mol Biomarkers 23: 759-765. doi: 10.1089/gtmb.2019.0092
|
| [157] |
Beaty TH, Taub MA, Scott AF, et al. (2013) Confirming genes influencing risk to cleft lip with/without cleft palate in a case-parent trio study. Hum Genet 132: 771-781. doi: 10.1007/s00439-013-1283-6
|
| [158] |
Moreno Uribe LM, Fomina T, Munger RG, et al. (2017) A Population-Based Study of Effects of Genetic Loci on Orofacial Clefts. J Dent Res 96: 1322-1329. doi: 10.1177/0022034517716914
|
| [159] |
Ghazali N, Rahman NA, Kannan TP, et al. (2015) Screening of Transforming Growth Factor Beta 3 and Jagged2 Genes in the Malay Population With Nonsyndromic Cleft Lip With or Without Cleft Palate. Cleft Palate Craniofac J 52: e88-94. doi: 10.1597/14-024
|
| [160] |
Simioni M, Araujo TK, Monlleo IL, et al. (2014) Investigation of genetic factors underlying typical orofacial clefts: mutational screening and copy number variation. J Hum Genet 60: 17-25. doi: 10.1038/jhg.2014.96
|
| [161] |
Shi M, Christensen K, Weinberg CR, et al. (2007) Orofacial cleft risk is increased with maternal smoking and specific detoxification-gene variants. Am J Hum Genet 80: 76-90. doi: 10.1086/510518
|
| [162] |
Song T, Wu D, Wang Y, et al. (2013) Association of NAT1 and NAT2 genes with nonsyndromic cleft lip and palate. Mol Med Rep 8: 211-216. doi: 10.3892/mmr.2013.1467
|
| [163] |
Sull JW, Liang KY, Hetmanski JB, et al. (2009) Maternal transmission effects of the PAX genes among cleft case-parent trios from four populations. Eur J Hum Genet 17: 831-839. doi: 10.1038/ejhg.2008.250
|
| [164] |
Laumonnier F, Holbert S, Ronce N, et al. (2005) Mutations in PHF8 are associated with X linked mental retardation and cleft lip/cleft palate. J Med Genet 42: 780-786. doi: 10.1136/jmg.2004.029439
|
| [165] |
Loenarz C, Ge W, Coleman ML, et al. (2010) PHF8, a gene associated with cleft lip/palate and mental retardation, encodes for an Nepsilon-dimethyl lysine demethylase. Hum Mol Genet 19: 217-222. doi: 10.1093/hmg/ddp480
|
| [166] |
Peanchitlertkajorn S, Cooper ME, Liu YE, et al. (2003) Chromosome 17: gene mapping studies of cleft lip with or without cleft palate in Chinese families. Cleft Palate Craniofac J 40: 71-79. doi: 10.1597/1545-1569_2003_040_0071_cgmsoc_2.0.co_2
|
| [167] |
Letra A, Silva RA, Menezes R, et al. (2007) MMP gene polymorphisms as contributors for cleft lip/palate: association with MMP3 but not MMP1. Arch Oral Biol 52: 954-960. doi: 10.1016/j.archoralbio.2007.04.005
|
| [168] |
Kumari P, Singh SK, Raman R (2019) TGFβ3, MSX1, and MMP3 as Candidates for NSCL±P in an Indian Population. Cleft Palate Craniofac J 56: 363-372. doi: 10.1177/1055665618775727
|
| [169] |
Letra A, Silva RM, Motta LG, et al. (2012) Association of MMP3 and TIMP2 promoter polymorphisms with nonsyndromic oral clefts. Birth Defects Res A Clin Mol Teratol 94: 540-548. doi: 10.1002/bdra.23026
|
| [170] |
Letra A, Zhao M, Silva RM, et al. (2014) Functional Significance of MMP3 and TIMP2 Polymorphisms in Cleft Lip/Palate. J Dent Res 93: 651-656. doi: 10.1177/0022034514534444
|
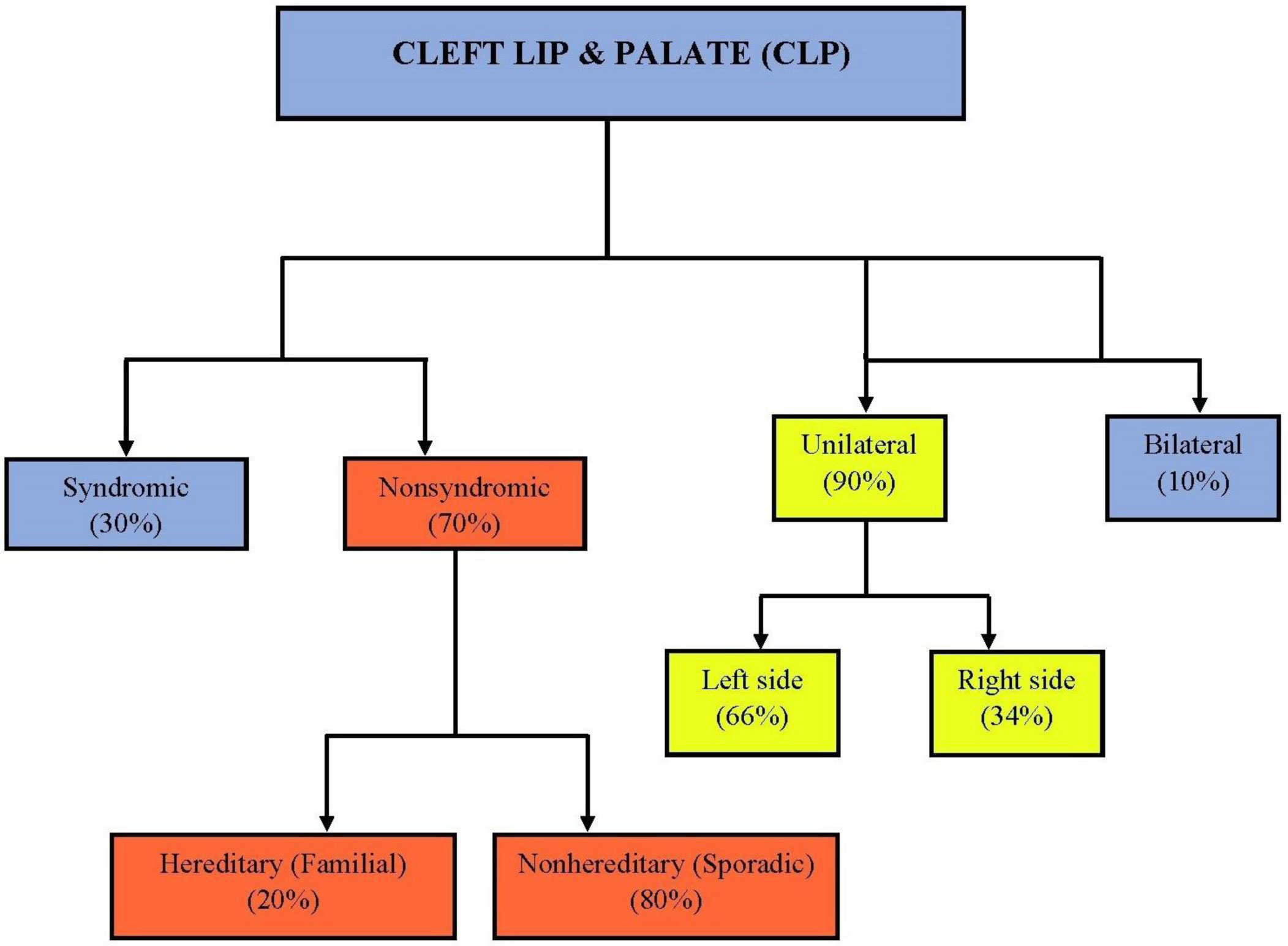
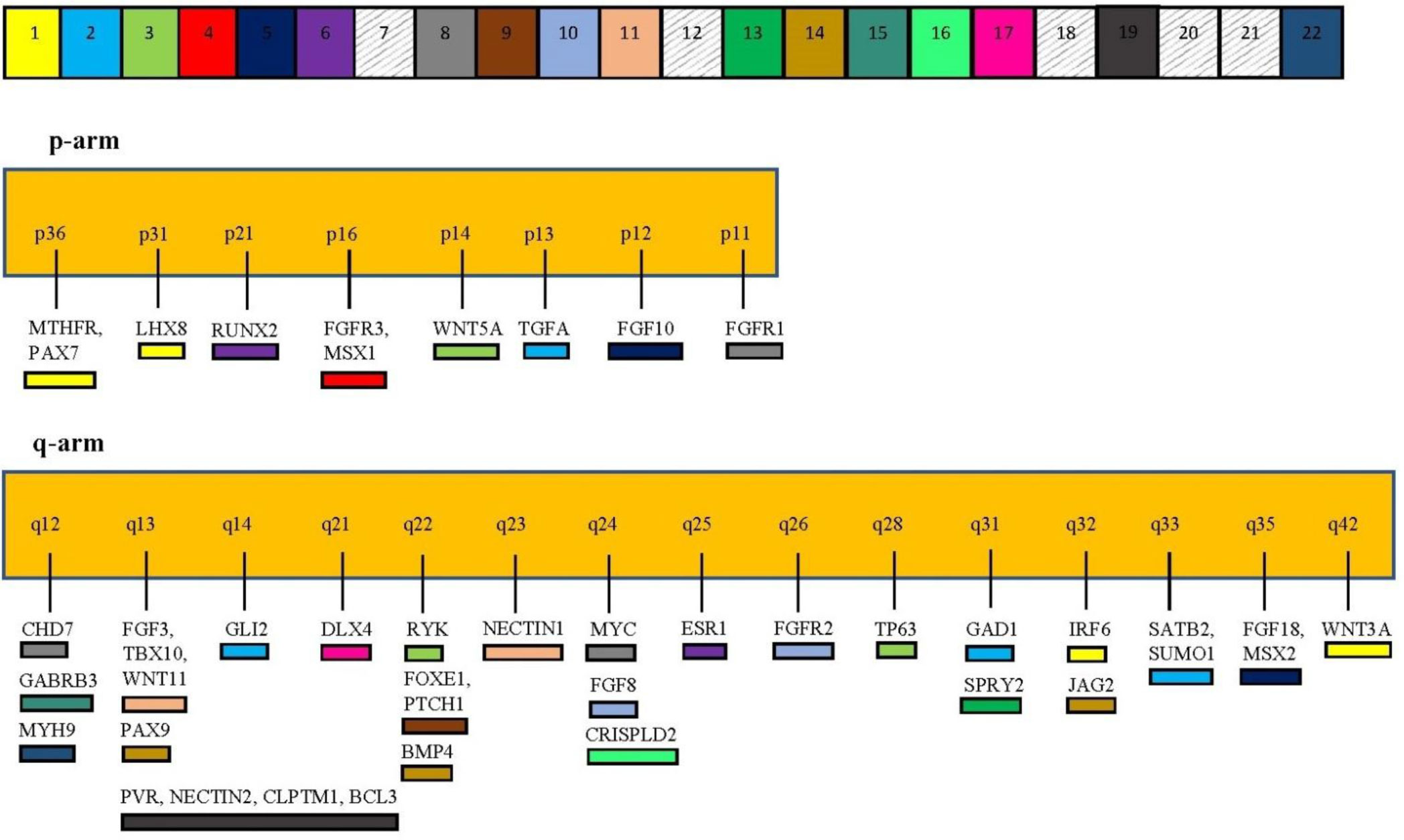
Figures(2) / Tables(1)

AN Mahamad Irfanulla Khan, CS Prashanth, N Srinath. Genetic etiology of cleft lip and cleft palate[J]. AIMS Molecular Science, 2020, 7(4): 328-348. doi: 10.3934/molsci.2020016







 DownLoad:
DownLoad: