Citation: Federica Martorana, Paolo Vigneri, Stefano Sergio Cordio, Concetta Martines, Giuseppe Novello, Marco Maria Aiello, Roberto Bordonaro, Hèctor J. Soto Parra. Efficacy of A Fluoropyrimidine plus Mitomycin C in Pretreated Patients with Metastatic Colorectal Cancer Eligible for Regorafenib: A Retrospective Study[J]. AIMS Medical Science, 2017, 4(4): 456-464. doi: 10.3934/medsci.2017.4.456

| [1] |
Ferlay J, Soerjomataram I, Dikshit R, et al. (2015) Cancer incidence and mortality worldwide: sources, methods and major patterns in GLOBOCAN 2012. Int J Cancer 136: E359-386. doi: 10.1002/ijc.29210

|
| [2] |
Siegel RL, Miller KD, Jemal A (2016) Cancer statistics. CA Cancer J Clin 66: 7-30. doi: 10.3322/caac.21332
|
| [3] |
Meta-analysis Group In C, Piedbois P, Rougier P, et al. (1998) Efficacy of intravenous continuous infusion of fluorouracil compared with bolus administration in advanced colorectal cancer. J Clin Oncol 16: 301-308. doi: 10.1200/JCO.1998.16.1.301
|
| [4] |
Kopetz S, Chang GJ, Overman MJ, et al. (2009) Improved survival in metastatic colorectal cancer is associated with adoption of hepatic resection and improved chemotherapy. J Clin Oncol 27: 3677-3683. doi: 10.1200/JCO.2008.20.5278
|
| [5] |
Waddell T, Cunningham D (2013) Evaluation of regorafenib in colorectal cancer and GIST. Lancet 381: 273-275. doi: 10.1016/S0140-6736(12)62006-6
|
| [6] | Vigneri PG, Tirro E, Pennisi MS, et al. (2015) The Insulin/IGF System in Colorectal Cancer Development and Resistance to Therapy. Front Oncol 5: 230. |
| [7] |
Chong G, Dickson JL, Cunningham D, et al. (2005) Capecitabine and mitomycin C as third-line therapy for patients with metastatic colorectal cancer resistant to fluorouracil and irinotecan. Br J Cancer 93: 510-514. doi: 10.1038/sj.bjc.6602733
|
| [8] | Rosati G, Rossi A, Germano D, et al. (2003) Raltitrexed and mitomycin-C as third-line chemotherapy for colorectal cancer after combination regimens including 5-fluorouracil, irinotecan and oxaliplatin: a phase II study. Anticancer Res 23: 2981-2985. |
| [9] | Lim DH, Park YS, Park BB, et al. (2005) Mitomycin-C and capecitabine as third-line chemotherapy in patients with advanced colorectal cancer: a phase II study. Cancer Chemother Pharmacol 56: 10-14. |
| [10] |
Kang EJ, Choi YJ, Kim JS, et al. (2010) Mitomycin-C, 5-fluorouracil, and leucovorin as a salvage therapy in patients with metastatic colorectal adenocarcinoma. Asia Pac J Clin Oncol 6: 286-291. doi: 10.1111/j.1743-7563.2010.01334.x
|
| [11] | Saif MW, Kaley K, Brennan M, et al. (2013) Mitomycin-C and capecitabine (MIXE) as salvage treatment in patients with refractory metastatic colorectal cancer: a retrospective study. Anticancer Res 33: 2743-2746. |
| [12] |
Zygulska AL, Krzemieniecki K (2015) Salvage chemotherapy in metastatic colorectal cancer with the combination of capecitabine and mitomycin C. Neoplasma 62: 793-797. doi: 10.4149/neo_2015_095
|
| [13] |
Dimou A, Syrigos KN, Saif MW (2010) Is there a role for mitomycin C in metastatic colorectal cancer? Expert Opin Investig Drugs 19: 723-735. doi: 10.1517/13543784.2010.485191
|
| [14] |
Ferrarotto R, Machado K, Mak MP, et al. (2012) A multicenter, multinational analysis of mitomycin C in refractory metastatic colorectal cancer. Eur J Cancer 48: 820-826. doi: 10.1016/j.ejca.2012.01.008
|
| [15] |
Wilhelm SM, Dumas J, Adnane L, et al. (2011) Regorafenib (BAY 73-4506): a new oral multikinase inhibitor of angiogenic, stromal and oncogenic receptor tyrosine kinases with potent preclinical antitumor activity. Int J Cancer 129: 245-255. doi: 10.1002/ijc.25864
|
| [16] |
Grothey A, Van Cutsem E, Sobrero A, et al. (2013) Regorafenib monotherapy for previously treated metastatic colorectal cancer (CORRECT): an international, multicentre, randomised, placebo-controlled, phase 3 trial. Lancet 381: 303-312. doi: 10.1016/S0140-6736(12)61900-X
|
| [17] | Jang GS, Kim MJ, Ha HI, et al. (2013) Comparison of RECIST version 1.0 and 1.1 in assessment of tumor response by computed tomography in advanced gastric cancer. Chin J Cancer Res 25: 689-694. |
| [18] |
Goldstein DA, Ahmad BB, Chen Q, et al. (2015) Cost-Effectiveness Analysis of Regorafenib for Metastatic Colorectal Cancer. J Clin Oncol 33: 3727-3732. doi: 10.1200/JCO.2015.61.9569
|
| [19] |
Kimura M, Usami E, Iwai M, et al. (2016) Comparison of cost-effectiveness of regorafenib and trifluridine/tipiracil combination tablet for treating advanced and recurrent colorectal cancer. Mol Clin Oncol 5: 635-640. doi: 10.3892/mco.2016.1020
|
| [20] |
Graziano F, Rulli E, Biagioli E, et al. (2016) Number needed to treat for pricing costly anticancer drugs: the example of regorafenib in metastatic colorectal cancer. Ann Oncol 27: 747-748. doi: 10.1093/annonc/mdw024
|
| [21] |
Mayer RJ, Van Cutsem E, Falcone A, et al. (2015) Randomized trial of TAS-102 for refractory metastatic colorectal cancer. N Engl J Med 372: 1909-1919. doi: 10.1056/NEJMoa1414325
|
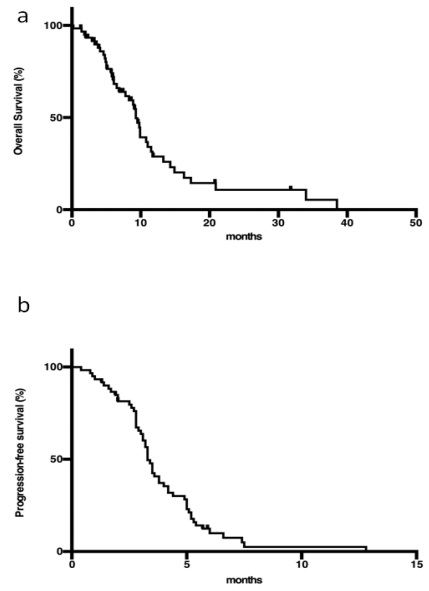
Figures(1) / Tables(1)

Federica Martorana, Paolo Vigneri, Stefano Sergio Cordio, Concetta Martines, Giuseppe Novello, Marco Maria Aiello, Roberto Bordonaro, Hèctor J. Soto Parra. Efficacy of A Fluoropyrimidine plus Mitomycin C in Pretreated Patients with Metastatic Colorectal Cancer Eligible for Regorafenib: A Retrospective Study[J]. AIMS Medical Science, 2017, 4(4): 456-464. doi: 10.3934/medsci.2017.4.456







 DownLoad:
DownLoad: