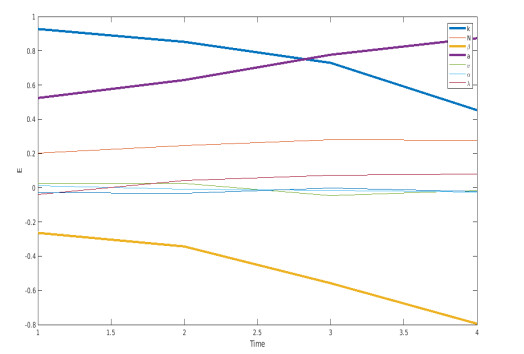
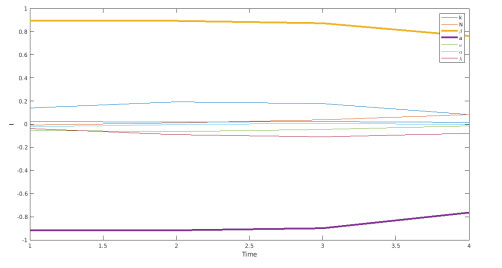
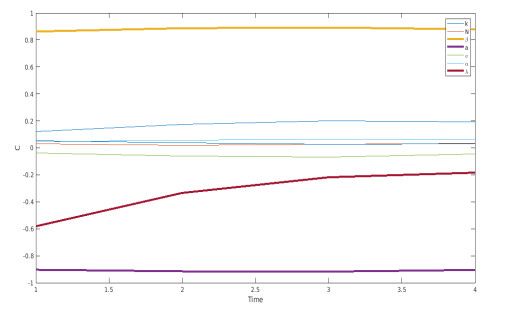
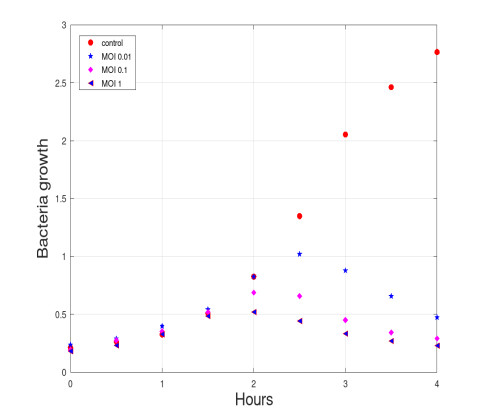
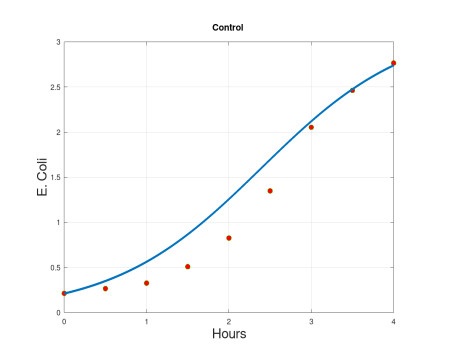
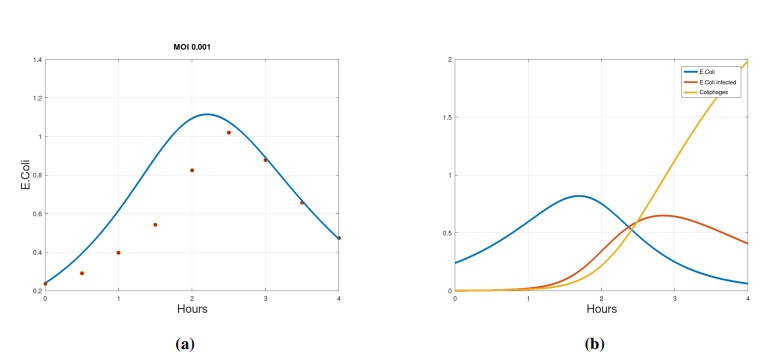
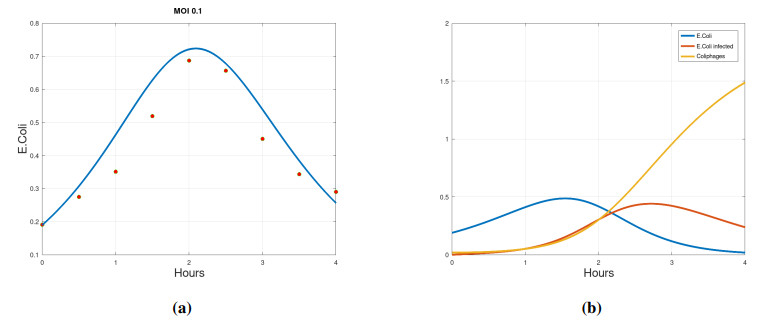
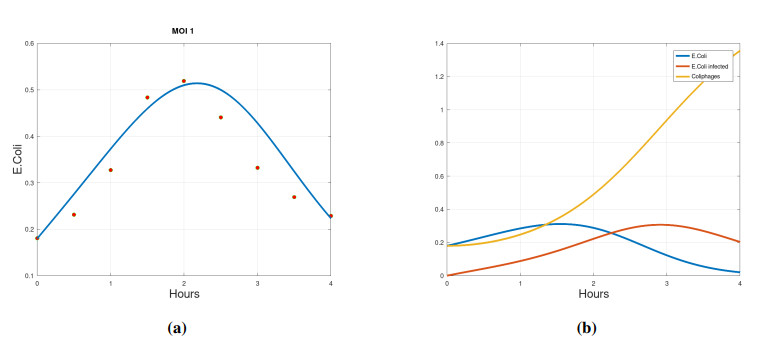
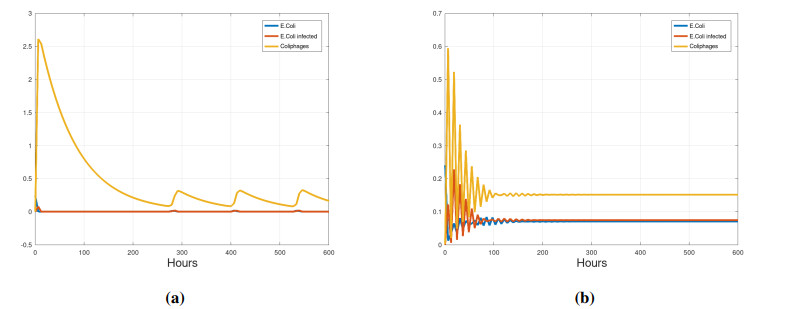
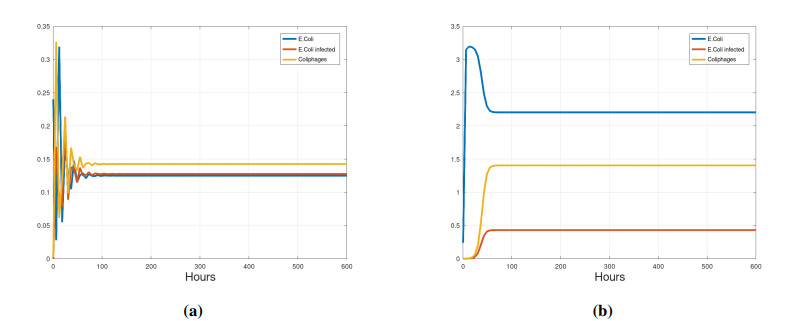
We propose a mathematical model based in ordinary differential equations between bacterial pathogen and Bacteriophages to describe the infection dynamics of these populations, for which we use a nonlinear function with an inhibitory effect. We study the stability of the model using the Lyapunov theory and the second additive compound matrix and perform a global sensitivity analysis to elucidate the most influential parameters in the model, besides we make a parameter estimation using growth data of Escherichia coli (E.coli) bacteria in presence of Coliphages (bacteriophages that infect E.coli) with different multiplicity of infection. We found a threshold that indicates whether the bacteriophage concentration will coexist with the bacterium (the coexistence equilibrium) or become extinct (phages extinction equilibrium), the first equilibrium is locally asymptotically stable while the other is globally asymptotically stable depending on the magnitude of this threshold. Beside we found that the dynamics of the model is particularly affected by infection rate of bacteria and Half-saturation phages density. Parameter estimation show that all multiplicities of infection are effective in eliminating infected bacteria but the smaller one leaves a higher number of bacteriophages at the end of this elimination.
Citation: Miller Cerón Gómez, Eduardo Ibarguen Mondragon, Eddy Lopez Molano, Arsenio Hidalgo-Troya, Maria A. Mármol-Martínez, Deisy Lorena Guerrero-Ceballos, Mario A. Pantoja, Camilo Paz-García, Jenny Gómez-Arrieta, Mariela Burbano-Rosero. Mathematical model of interaction Escherichia coli and Coliphages[J]. Mathematical Biosciences and Engineering, 2023, 20(6): 9712-9727. doi: 10.3934/mbe.2023426
We propose a mathematical model based in ordinary differential equations between bacterial pathogen and Bacteriophages to describe the infection dynamics of these populations, for which we use a nonlinear function with an inhibitory effect. We study the stability of the model using the Lyapunov theory and the second additive compound matrix and perform a global sensitivity analysis to elucidate the most influential parameters in the model, besides we make a parameter estimation using growth data of Escherichia coli (E.coli) bacteria in presence of Coliphages (bacteriophages that infect E.coli) with different multiplicity of infection. We found a threshold that indicates whether the bacteriophage concentration will coexist with the bacterium (the coexistence equilibrium) or become extinct (phages extinction equilibrium), the first equilibrium is locally asymptotically stable while the other is globally asymptotically stable depending on the magnitude of this threshold. Beside we found that the dynamics of the model is particularly affected by infection rate of bacteria and Half-saturation phages density. Parameter estimation show that all multiplicities of infection are effective in eliminating infected bacteria but the smaller one leaves a higher number of bacteriophages at the end of this elimination.

| [1] |
A. M. Comeau, G. F. Hatfull, H. M. Krisch, D. Lindell, N. H. Mann, D. Prangishvili, Exploring the prokaryotic virosphere, Res. Microbiol., 159 (2008), 306–313. https://doi.org/10.1016/j.resmic.2008.05.001 doi: 10.1016/j.resmic.2008.05.001

|
| [2] |
M. R. Clokie, A. D. Millard, A. V. Letarov, S. Heaphy, Phages in nature, Bacteriophage, 1 (2011), 31–45. https://doi.org/10.4161/bact.1.1.14942 doi: 10.4161/bact.1.1.14942
|
| [3] |
C. Howard-Varona, K. R. Hargreaves, S. T. Abedon, M. B. Sullivan, Lysogeny in nature: mechanisms, impact and ecology of temperate phages, ISME J., 11 (2017), 1511–1520. https://doi.org/10.1038/ismej.2017.16 doi: 10.1038/ismej.2017.16
|
| [4] |
J. R. Clark, J. B. March, Bacteriophages and biotechnology: vaccines, gene therapy and antibacterials, Trends Biotechnol., 24 (2006), 212–218. https://doi.org/10.1016/j.tibtech.2006.03.003 doi: 10.1016/j.tibtech.2006.03.003
|
| [5] |
I. U. Haq, W. N. Chaudhry, M. N. Akhtar, S. Andleeb, I. Qadri, Bacteriophages and their implications on future biotechnology: a review, Virol. J., 9 (2012), 1–8. https://doi.org/10.1186/1743-422X-9-9 doi: 10.1186/1743-422X-9-9
|
| [6] |
C. Loc-Carrillo, S. T. Abedon, Pros and cons of phage therapy, Bacteriophage, 1 (2011), 111–114. https://doi.org/10.4161/bact.1.2.14590 doi: 10.4161/bact.1.2.14590
|
| [7] |
R. Jain, A. L. Knorr, J. Bernacki, R. Srivastava, Investigation of bacteriophage MS2 viral dynamics using model discrimination analysis and the implications for phage therapy, Biotechnol. Progr., 22 (2006), 1650–1658. https://doi.org/10.1021/bp060161s doi: 10.1021/bp060161s
|
| [8] |
G. Beke, M. Stano, L. Klucar, Modelling the interaction between bacteriophages and their bacterial hosts, Math. Biosci., 279 (2016), 27–32. https://doi.org/10.1016/j.mbs.2016.06.009 doi: 10.1016/j.mbs.2016.06.009
|
| [9] |
B. J. Cairns, A. R. Timms, V. A. Jansen, I. F. Connerton, R. J. Payne, Quantitative models of in vitro bacteriophage–host dynamics and their application to phage therapy, PLoS Pathog., 5 (2009), e1000253. https://doi.org/10.1371/journal.ppat.1000253 doi: 10.1371/journal.ppat.1000253
|
| [10] |
H. Ndongmo Teytsa, B. Tsanou, S. Bowong, J. M. Lubuma, Bifurcation analysis of a phage-bacteria interaction model with prophage induction, Math. Med. Biol., 38 (2021), 28–58. https://doi.org/10.1093/imammb/dqaa010 doi: 10.1093/imammb/dqaa010
|
| [11] |
X. Li, R. Huang, M. He, Dynamics model analysis of bacteriophage infection of bacteria, Adv. Differ. Equations, 2021 (2021), 1–11. https://doi.org/10.1186/s13662-021-03466-x doi: 10.1186/s13662-021-03466-x
|
| [12] |
S. Pagliarini, A. Korobeinikov, A mathematical model of marine bacteriophage evolution, Roy. Soc. Open Sci., 5 (2018), 171661. https://doi.org/10.1098/rsos.171661 doi: 10.1098/rsos.171661
|
| [13] |
C. C. McCluskey, P. van den Driessche, Global analysis of two tuberculosis models, J. Dyn. Differ. Equations, 16 (2004), 139–166. https://doi.org/10.1023/B:JODY.0000041283.66784.3e doi: 10.1023/B:JODY.0000041283.66784.3e
|
| [14] | M. Y. Li, J. S. Muldowney, On RA smith's autonomous convergence theorem, Rocky Mount. J. Math., 25 (1995), 365–379. |
| [15] |
S. Marino, I. B. Hogue, C. J. Ray, D. E. Kirschner, A methodology for performing global uncertainty and sensitivity analysis in systems biology, J. Theor. Biol., 254 (2008), 178–196. https://doi.org/10.1016/j.jtbi.2008.04.011 doi: 10.1016/j.jtbi.2008.04.011
|
| [16] |
M. Konopacki, B. Grygorcewicz, B. Dolegowska, M. Kordas, R. Rakoczy, Phagescore: A simple method for comparative evaluation of bacteriophages lytic activity, Biochem. Eng. J., 161 (2020), 107652. https://doi.org/10.1016/j.bej.2020.107652 doi: 10.1016/j.bej.2020.107652
|
| [17] |
M. C. Gómez, H. M. Yang, Mathematical model of the immune response to dengue virus, J. Appl. Math. Comput., 63 (2020), 455–478. https://doi.org/10.1007/s12190-020-01325-8 doi: 10.1007/s12190-020-01325-8
|
| [18] |
N. Principi, E. Silvestri, S. Esposito, Advantages and limitations of bacteriophages for the treatment of bacterial infections, Front. pharmacol., 10 (2019), 513. https://doi.org/10.3389/fphar.2019.00513 doi: 10.3389/fphar.2019.00513
|
| [19] |
M. Merabishvili, C. Vervaet, J. P. Pirnay, D. De Vos, G. Verbeken, J. Mast, et al., Stability of Staphylococcus aureus phage ISP after freeze-drying (lyophilization). PloS One, 8 (2013), e68797. https://doi.org/10.1371/journal.pone.0068797 doi: 10.1371/journal.pone.0068797
|
Figures(10) / Tables(2)

Miller Cerón Gómez, Eduardo Ibarguen Mondragon, Eddy Lopez Molano, Arsenio Hidalgo-Troya, Maria A. Mármol-Martínez, Deisy Lorena Guerrero-Ceballos, Mario A. Pantoja, Camilo Paz-García, Jenny Gómez-Arrieta, Mariela Burbano-Rosero. Mathematical model of interaction Escherichia coli and Coliphages[J]. Mathematical Biosciences and Engineering, 2023, 20(6): 9712-9727. doi: 10.3934/mbe.2023426







 DownLoad:
DownLoad: