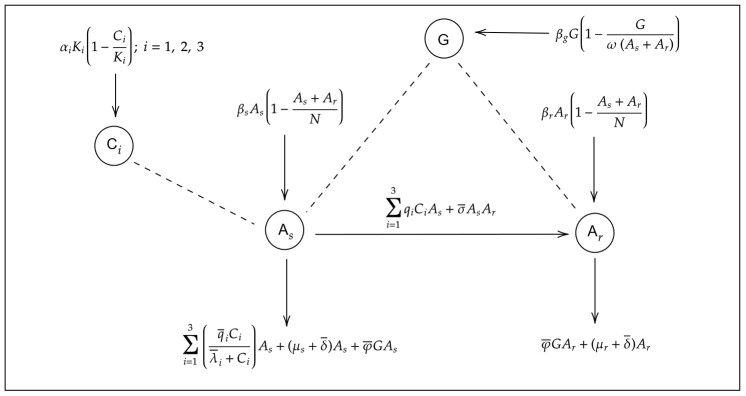
In this work, we formulated a mathematical model to describe growth, acquisition of bacterial resistance, and immune response for Helicobacter pylori (H. pylori). The qualitative analysis revealed the existence of five equilibrium solutions: (ⅰ) An infection-free state, in which the bacterial population and immune cells are suppressed, (ⅱ) an endemic state only with resistant bacteria without immune cells, (ⅲ) an endemic state only with resistant bacteria and immune cells, (ⅳ) an endemic state of bacterial coexistence without immune cells, and (ⅴ) an endemic coexistence state with immune response. The stability analysis showed that the equilibrium solutions (ⅰ) and (ⅳ) are locally asymptotically stable, whereas the equilibria (ⅱ) and (ⅲ) are unstable. We found four threshold conditions that establish the existence and stability of equilibria, which determine when the populations of sensitive H. pylori and resistant H. pylori are controlled or eliminated, or when the infection progresses only with resistant bacteria or with both bacterial populations. The numerical simulations corroborated the qualitative analysis, and provided information on the emergence of a limit cycle that breaks the stability of the coexistence equilibrium. The results revealed that the key to controlling bacterial progression is to keep bacterial growth thresholds below 1; this can be achieved by applying an appropriate combination of antibiotics and correct stimulation of the immune response. Otherwise, when bacterial growth thresholds exceed 1, the bacterial persistence scenarios mentioned above occur.
Citation: Edgar Alberto Vega Noguera, Simeón Casanova Trujillo, Eduardo Ibargüen-Mondragón. A within-host model on the interactions of sensitive and resistant Helicobacter pylori to antibiotic therapy considering immune response[J]. Mathematical Biosciences and Engineering, 2025, 22(1): 185-224. doi: 10.3934/mbe.2025009
In this work, we formulated a mathematical model to describe growth, acquisition of bacterial resistance, and immune response for Helicobacter pylori (H. pylori). The qualitative analysis revealed the existence of five equilibrium solutions: (ⅰ) An infection-free state, in which the bacterial population and immune cells are suppressed, (ⅱ) an endemic state only with resistant bacteria without immune cells, (ⅲ) an endemic state only with resistant bacteria and immune cells, (ⅳ) an endemic state of bacterial coexistence without immune cells, and (ⅴ) an endemic coexistence state with immune response. The stability analysis showed that the equilibrium solutions (ⅰ) and (ⅳ) are locally asymptotically stable, whereas the equilibria (ⅱ) and (ⅲ) are unstable. We found four threshold conditions that establish the existence and stability of equilibria, which determine when the populations of sensitive H. pylori and resistant H. pylori are controlled or eliminated, or when the infection progresses only with resistant bacteria or with both bacterial populations. The numerical simulations corroborated the qualitative analysis, and provided information on the emergence of a limit cycle that breaks the stability of the coexistence equilibrium. The results revealed that the key to controlling bacterial progression is to keep bacterial growth thresholds below 1; this can be achieved by applying an appropriate combination of antibiotics and correct stimulation of the immune response. Otherwise, when bacterial growth thresholds exceed 1, the bacterial persistence scenarios mentioned above occur.

| [1] | E. C. García, R. García-González, Helicobacter pylori y la respuesta inmune, Rev. Latinoam. Patol. Clin. Med. Lab., 62 (2015), 112–118. |
| [2] |
J. K. Y. Hooi, W. Y. Lai, W. K. Ng, M. M. Y. Suen, Global prevalence of Helicobacter pylori infection: systematic review and meta-analysis, Gastroenterology, 153 (2017), 420–429. https://doi.org/10.1053/j.gastro.2017.04.022 doi: 10.1053/j.gastro.2017.04.022

|
| [3] |
E. Tshibangu-Kabamba, Y. Yamaoka, Helicobacter pylori infection and antibiotic resistance from biology to clinical implications, Nat. Rev. Gastroenterol. Hepatol., 18 (2021), 613–629. https://doi.org/10.1038/s41575-021-00449-x doi: 10.1038/s41575-021-00449-x
|
| [4] |
A. Savoldi, E. Carrara, D. Y. Graham, M. Conti, E. Tacconelli, Prevalence of antibiotic resistance in Helicobacter pylori: A systematic review and Meta-analysis in World Health Organization Regions, Gastroenterology, 155 (2018), 1372–1382. https://doi.org/10.1053/j.gastro.2018.07.007 doi: 10.1053/j.gastro.2018.07.007
|
| [5] |
F. Megraud, TH pylori antibiotic resistance: prevalence, importance, and advances in testing, Gut, 53 (2004), 1374–1384. https://doi.org/10.1136/gut.2003.022111 doi: 10.1136/gut.2003.022111
|
| [6] |
I. M. Joseph, Y. Zavros, J. L. Merchant, D. Kirschner, A model for integrative study of human gastric acid secretion, J. Appl. Physiol., 94 (2003), 1602–1618. https://doi.org10.1152/japplphysiol.00281.2002 doi: 10.1152/japplphysiol.00281.2002
|
| [7] |
S. Marino, The importance of an inter-compartmental delay in a model for human gastric acid secretion, Bull. Math. Biol., 65 (2003), 963–990. https://doi.org/10.1016/s0092-8240(03)00046-6 doi: 10.1016/s0092-8240(03)00046-6
|
| [8] |
D. Sud, I. M. Joseph, D. Kirschner, Predicting eficacy of proton pump inhibitors in regulating gastric acid secretion, J. Biol. Syst., 12 (2004), 1–34. https://doi.org/10.1142/s0218339004000999 doi: 10.1142/s0218339004000999
|
| [9] |
M. J. Blaser, D. E. Kirschner, Dynamics of Helicobacter pylori colonization in relation to the host response, Proc. Natl. Acad. Sci. U.S.A., 96 (1999), 8359–8364. https://doi.org/10.1073/pnas.96.15.8359 doi: 10.1073/pnas.96.15.8359
|
| [10] |
I. M. Joseph, D. Kirschner, A model for the study of Helicobacter pylori interaction with human gastric acid secretion, J. Theor. Biol., 228 (2004), 55–80. https://doi.org/10.1016/j.jtbi.2003.12.004 doi: 10.1016/j.jtbi.2003.12.004
|
| [11] |
D. E. Kirschner, M. J. Blaser, The dynamics of Helicobacter pylori infection of the human stomach, J. Theor. Biol., 176 (1995), 281–290. https://doi.org/10.1006/jtbi.1995.0198 doi: 10.1006/jtbi.1995.0198
|
| [12] |
E. Ibargüen-Mondragón, S. Mosquera, M. Cerón, E. M. Burbano-Rosero, S. P. Hidalgo-Bonilla, L. Esteva, et al., Mathematical modeling on bacterial resistance to multiple antibiotics caused by spontaneous mutations, BioSystems, 117 (2014), 60–67. https://doi.org/10.1016/j.biosystems.2014.01.005 doi: 10.1016/j.biosystems.2014.01.005
|
| [13] | J. Romero, E. Ibargüen-Mondragón, Sobre la resistencia bacteriana hacia antibióticos de acción bactericida y bacteriostática, Rev. Integración, 32 (2014), 101–116. |
| [14] |
E. Ibargüen-Mondragón, J. P. Romero-Leiton, L. Esteva, E. M. Burbano-Rosero, Mathematical modeling of bacterial resistance to antibiotics by mutations and plasmids, J. Biol. Syst., 24 (2016), 129–146. https://doi.org/10.1142/s0218339016500078 doi: 10.1142/s0218339016500078
|
| [15] |
E. Ibargüen-Mondragón, J. Romero, P. Jhoana, L. Lourdes, M. Gómez Cerón, S. P. Hidalgo-Bonilla, Stability and periodic solutions for a model of bacterial resistance to antibiotics caused by mutations and plasmids, Appl. Math. Model., 76 (2019), 238–251. https://doi.org/10.1016/j.apm.2019.06.017 doi: 10.1016/j.apm.2019.06.017
|
| [16] |
E. Ibargüen-Mondragón, M. C. Gómez, E. M. Burbano-Rosero, Assessing the role of bacterial plasmid replication in a competition model of sensitive and resistant bacteria to antibiotics, AIMS Math., 6 (2021), 9446–9467. https://doi.org/10.3934/math.2021549 doi: 10.3934/math.2021549
|
| [17] |
E. Ibargüen-Mondragón, K. Prieto, S. P. Hidalgo-Bonilla, A model on bacterial resistance considering a generalized law of mass action for plasmid replication, J. Biol. Syst., 29 (2021), 375–412. https://doi.org/10.1142/s0218339021400118 doi: 10.1142/s0218339021400118
|
| [18] |
B. Daşbaşı, İ. Öztürk, Mathematical modelling of bacterial resistance to multiple antibiotics and immune system response, SpringerPlus, 5 (2016), 1–17. https://doi.org/10.1186/s40064-016-2017-8 doi: 10.1186/s40064-016-2017-8
|
| [19] |
B. Daşbaşı, İ. Öztürk, On the stability analysis of the general mathematical modeling of bacterial infection, Int. J. Eng. Appl. Sci., 10 (2018), 93–117. https://doi.org/10.24107/ijeas.445520 doi: 10.24107/ijeas.445520
|
| [20] |
S. Arenillas, A. Godoy, H. Einisman, D. García, P. Harris, Regulación de la respuesta inmune frente a la infección por Helicobacter pylori, Rev. Chil. Pediatr., 73 (2003), 108–115. https://doi.org/10.4067/s0370-41062002000200002 doi: 10.4067/s0370-41062002000200002
|
| [21] |
B. Bjorkholm, M. Sjolund, Mutation frequency and biological cost of antibiotic resistance in Helicobacter pylori, Proc. Natl. Acad. Sci. U.S.A., 98 (2001), 14607–14612. https://doi.org/10.1073/pnas.241517298 doi: 10.1073/pnas.241517298
|
| [22] |
A. Hanafi, W. Lee, M. F. Loke, X. Teh, A. Shaari, M. Dinarvand, et al., Molecular and proteomic analysis of levofloxacin and metronidazole resistant Helicobacter pylori, Front. Microbiol., 7 (2016), 1–12. https://doi.org/10.3389/fmicb.2016.02015 doi: 10.3389/fmicb.2016.02015
|
| [23] |
A. Handel, R. Regoes, R. Antia, The role of compensatory mutations in the emergence of drug resistance, PLoS Comput. Biol., 2 (2006), 1262–1270. https://doi.org/10.1371/journal.pcbi.0020137 doi: 10.1371/journal.pcbi.0020137
|
| [24] |
D. J. Austin, R. M. Anderson, Studies of antibiotic resistance within the patient, hospitals and the community using simple mathematical models, Philos. Trans. R. Soc. London, Ser. B, 354 (1999), 721–738. https://doi.org/10.1098/rstb.1999.0425 doi: 10.1098/rstb.1999.0425
|
| [25] |
M. Jacobs, N. Grégoire, W. Couet, J. B. Bulitta, Distinguishing antimicrobial models with different resistance mechanisms via population pharmacodynamic modeling, PLoS Comput. Biol., 12 (2016), 1–19. https://doi.org/10.1371/journal.pcbi.1004782 doi: 10.1371/journal.pcbi.1004782
|
| [26] |
M. Lipsitch, B. R. Levin, The population dynamics of antimicrobial chemotherapy, Antimicrob. Agents Chemother., 41 (1997), 363–373. https://doi.org/10.1128/aac.41.2.363 doi: 10.1128/aac.41.2.363
|
| [27] |
M. Nikolaou, V. H. Tam, A new modeling approach to the effect of antimicrobial agents on heterogeneous microbial populations, J. Math. Biol., 52 (2006), 154–182. https://doi.org/10.1007/s00285-005-0350-6 doi: 10.1007/s00285-005-0350-6
|
| [28] |
K. Paterson, A. Hoyle, G. Ochoa, C. Baker-Austin, N. G. Taylor, Optimising antibiotic usage to treat bacterial infections, Sci. Rep., 6 (2016), 1–10. https://doi.org/10.1038/srep37853 doi: 10.1038/srep37853
|
| [29] | P. Macheras, A. Iliadis, Modeling in Biopharmaceutics, Pharmacokinetics, and Pharmacodynamics: Homogeneous and Heterogeneous Approaches, Springer Cham, New York, 2006. http://doi.org/10.1007/978-3-319-27598-7 |
| [30] | W. Otero, M. Gómez, L. Otero, A. Trespalacios, Helicobacter pylori: cómo se trata en el 2018?, Rev. Gastroenterol. Peru., 38 (2018), 54–63. |
| [31] |
A. Handel, E. Margolis, B. R. Levin, Exploring the role of the immune response in preventing antibiotic resistance, J. Theor. Biol., 256 (2008), 655–662. https://doi.org/10.1016/j.jtbi.2008.10.025 doi: 10.1016/j.jtbi.2008.10.025
|
| [32] |
G. E. Wang, J. M. Trevor, Q. Jiang, D. E. Taylor, Spontaneous mutations that confer antibiotic resistance in Helicobacter pylori, Antimicrob. Agents Chemother., 45 (2001), 727–733. https://doi.org/10.1128/aac.45.3.727-733.2001 doi: 10.1128/aac.45.3.727-733.2001
|
| [33] |
J. Faghri, F. Poursina, S. Moghim, H. Z. Esfahani, B. N. Esfahani, H. Fazeli, et al., Morphological and bactericidal effects of different antibiotics on Helicobacter pylori, Jundishapur J. Microbiol., 7 (2014), 1–6. https://doi.org/10.5812/jjm.8704 doi: 10.5812/jjm.8704
|
| [34] |
S. Fujimura, S. Kato, T. Kawamura, A. Watanabe, In vitro activity of rifampicin against Helicobacter pylori isolated from children and adults, J. Antimicrob. Chemother., 49 (2002), 541–543. https://doi.org/10.1093/jac/49.3.541 doi: 10.1093/jac/49.3.541
|
| [35] |
E. Begg, R. Robson, D. A. Saunders, G. G. Graham, R. C. Buttimore, A. M. Neill, et al., The pharmacokinetics of oral fleroxacin and ciprofloxacin in plasma and sputum during acute and chronic dosing, Br. J. Clin. Pharmacol., 49 (2000), 32–38. https://doi.org/10.1046/j.1365-2125.2000.00105.x doi: 10.1046/j.1365-2125.2000.00105.x
|
| [36] |
D. Mainz, K. Borner, P. Koeppe, J. Kotwas, H. Lode, Pharmacokinetics of lansoprazole, amoxicillin and clarithromycin after simultaneous and single administration, J. Antimicrob. Chemother., 50 (2002), 699–706. https://doi.org/10.1093/jac/dkf172 doi: 10.1093/jac/dkf172
|
| [37] | R. Mejías-Luque, M. Gerhard, Immune Evasion Strategies and Persistence of Helicobacter Pylori, Springer International Publishing, New York, 2017. https://doi.org/10.1007/978-3-319-50520-63 |
| [38] |
A. Karkhah, S. Ebrahimpour, M. Rostamtabar, V. Koppolu, S. Darvish, V. K. R. Vasigala, et al., Helicobacter pylori evasion strategies of the host innate and adaptive immune responses to survive and develop gastrointestinal diseases, Microbiol. Res., 218 (2019), 49–57. https://doi.org/10.1016/j.micres.2018.09.011 doi: 10.1016/j.micres.2018.09.011
|










Figures(11) / Tables(1)

Edgar Alberto Vega Noguera, Simeón Casanova Trujillo, Eduardo Ibargüen-Mondragón. A within-host model on the interactions of sensitive and resistant Helicobacter pylori to antibiotic therapy considering immune response[J]. Mathematical Biosciences and Engineering, 2025, 22(1): 185-224. doi: 10.3934/mbe.2025009







 DownLoad:
DownLoad: