Citation: A. M. Elaiw, N. H. AlShamrani, A. D. Hobiny. Stability of an adaptive immunity delayed HIV infection model with active and silent cell-to-cell spread[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 6401-6458. doi: 10.3934/mbe.2020337

| [1] |
M. A. Nowak, C. R. M. Bangham, Population dynamics of immune responses to persistent viruses, Science, 272 (1996), 74-79. doi: 10.1126/science.272.5258.74

|
| [2] | A. M. Elaiw, R. M. Abukwaik, E. O. Alzahrani, Global properties of a cell mediated immunity in HIV infection model with two classes of target cells and distributed delays, Int. J. Biomath., 7 (2014), Article ID 1450055. |
| [3] | W. Chen, N. Tuerxun, Z. Teng, The global dynamics in a wild-type and drug-resistant HIV infection model with saturated incidence, Adv. Differ. Equ., 2020 (2020), Article Number: 25. |
| [4] |
A. M. Elaiw, S. A. Azoz, Global properties of a class of HIV infection models with BeddingtonDeAngelis functional response, Math. Methods Appl. Sci., 36 (2013), 383-394. doi: 10.1002/mma.2596
|
| [5] |
X. Zhou, L. Zhang, T. Zheng, H. Li, Z. Teng, Global stability for a class of HIV virus-to-cell dynamical model with Beddington-DeAngelis functional response and distributed time delay, Math. Biosci. Eng., 17 (2020), 4527-4543. doi: 10.3934/mbe.2020250
|
| [6] |
G. Huang, Y. Takeuchi, W. Ma, Lyapunov functionals for delay differential equations model of viral infections, SIAM J. Appl. Math., 70 (2010), 2693-2708. doi: 10.1137/090780821
|
| [7] |
A. M. Elaiw, S. F. Alshehaiween, Global stability of delay-distributed viral infection model with two modes of viral transmission and B-cell impairment, Math. Methods Appl. Sci., 43 (2020), 6677-6701. doi: 10.1002/mma.6408
|
| [8] |
A. M. Elaiw, A. A. Raezah, Stability of general virus dynamics models with both cellular and viral infections and delays, Math. Methods Appl. Sci., 40 (2017), 5863-5880. doi: 10.1002/mma.4436
|
| [9] |
A. M. Elaiw, M. A. Alshaikh, Stability analysis of a general discrete-time pathogen infection model with humoral immunity, J. Differ. Equ. Appl., 25 (2019), 1149-1172. doi: 10.1080/10236198.2019.1662411
|
| [10] |
D. Li, W. Ma, Asymptotic properties of a HIV-1 infection model with time delay, J. Math. Anal. Appl., 335 (2007), 683-691. doi: 10.1016/j.jmaa.2007.02.006
|
| [11] |
R. V. Culshaw, S. Ruan, A delay-differential equation model of HIV infection of CD4+ T-cells, Math. Biosci., 165 (2000), 27-39. doi: 10.1016/S0025-5564(00)00006-7
|
| [12] | A. M. Elaiw, M. A. Alshaikh, Stability of a discrete-time general delayed viral model with antibody and cell-mediated immune responses, Adv. Differ. Equ., 2020 (2020), Article Number: 54. |
| [13] |
H. Kong, G. Zhang, K. Wang, Stability and Hopf bifurcation in a virus model with selfproliferation and delayed activation of immune cells, Math. Biosci., 17 (2020), 4384-4405. doi: 10.3934/mbe.2020242
|
| [14] |
H. Shu, L. Wang, J. Watmough, Global stability of a nonlinear viral infection model with infinitely distributed intracellular delays and CTL imune responses, SIAM J. Appl. Math., 73 (2013), 1280-1302. doi: 10.1137/120896463
|
| [15] |
J. Wang, C. Qin, Y. Chen, X. Wang, Hopf bifurcation in a CTL-inclusive HIV-1 infection model with two time delays, Math. Biosci. Eng., 16 (2019), 2587-2612. doi: 10.3934/mbe.2019130
|
| [16] | T. Kajiwara, T. Sasaki, A note on the stability analysis of pathogen-immune interaction dynamics, Discrete Contin. Dyn. Syst. Ser. B, 4 (2004), 615-622. |
| [17] | K. Hattaf, Global stability and Hopf bifurcation of a generalized viral infection model with multidelays and humoral immunity, Phys. A, 545 (2020), Article ID 123689. |
| [18] |
A. Murase, T. Sasaki, T. Kajiwara, Stability analysis of pathogen-immune interaction dynamics, J. Math. Biol., 51 (2005), 247-267. doi: 10.1007/s00285-005-0321-y
|
| [19] |
D. Wodarz, Hepatitis C virus dynamics and pathology: The role of CTL and antibody responses, J. Gen. Virol., 84 (2003), 1743-1750. doi: 10.1099/vir.0.19118-0
|
| [20] |
P. Dubey, U. S. Dubey, B. Dubey, Modeling the role of acquired immune response and antiretroviral therapy in the dynamics of HIV infection, Math. Comput. Simul., 144 (2018), 120-137. doi: 10.1016/j.matcom.2017.07.006
|
| [21] |
Y. Su, D. Sun, L. Zhao, Global analysis of a humoral and cellular immunity virus dynamics model with the Beddington-DeAngelis incidence rate, Math. Methods Appl. Sci., 38 (2015), 2984-2993. doi: 10.1002/mma.3274
|
| [22] |
N. Yousfi, K. Hattaf, A. Tridane, Modeling the adaptive immune response in HBV infection, J. Math. Biol., 63 (2011), 933-957. doi: 10.1007/s00285-010-0397-x
|
| [23] | A. Perelson, A. Neumann, M. Markowitz, J. Leonard, D. Ho, HIV-1 dynamics in vivo: virion clearance rate, infected cell life-span, and viral generation time, Science, 271 (1996), 1582-1586. |
| [24] | Y. Yan, W. Wang, Global stability of a five-dimensional model with immune responses and delay, Discrete Contin. Dyn. Syst. Ser. B, 17 (2012), 401-416. |
| [25] |
X. Wang, S. Liu, A class of delayed viral models with saturation infection rate and immune response, Math. Methods Appl. Sci., 36 (2013), 125-142. doi: 10.1002/mma.2576
|
| [26] | J. Wang, J. Pang, T. Kuniya, Y. Enatsu, Global threshold dynamics in a five-dimensional virus model with cell-mediated, humoral immune responses and distributed delays, Appl. Math. Comput., 241 (2014), 298-316. |
| [27] |
A. M. Elaiw, N. H. AlShamrani, Stability of a general delay-distributed virus dynamics model with multi-staged infected progression and immune response, Math. Methods Appl. Sci., 40 (2017), 699-719. doi: 10.1002/mma.4002
|
| [28] |
C. Jolly, Q. Sattentau, Retroviral spread by induction of virological synapses, Traffic, 5 (2004), 643-650. doi: 10.1111/j.1600-0854.2004.00209.x
|
| [29] | S. Iwami, J. S. Takeuchi, S. Nakaoka, F. Mammano, F. Clavel, H. Inaba, et al., Cell-to-cell infection by HIV contributes over half of virus infection, Elife, 4 (2015), e08150. |
| [30] |
N. L. Komarova, D. Wodarz, Virus dynamics in the presence of synaptic transmission, Math. Biosci., 242 (2013), 161-171. doi: 10.1016/j.mbs.2013.01.003
|
| [31] | A. Sigal, J. T. Kim, A. B. Balazs, E. Dekel, A. Mayo, R. Milo, et al., Cell-to-cell spread of HIV permits ongoing replication despite antiretroviral therapy, Nature, 477 (2011), 95-98. |
| [32] | T. Guo, Z. Qiu, L. Rong, Analysis of an HIV model with immune responses and cell-to-cell transmission, Bull. Malays. Math. Sci. Soc., 43 (2018), 581-607. |
| [33] | J. Lin, R. Xu, X. Tian, Threshold dynamics of an HIV-1 model with both viral and cellular infections, cell-mediated and humoral immune responses, Math. Biosci. Eng., 16 (2018), 292-319. |
| [34] | K. Hattaf, N. Yousfi, Modeling the adaptive immunity and both modes of transmission in HIV infection, Computation, 6 (2018), Article ID 37. |
| [35] | T.-W. Chun, L. Stuyver, S. B. Mizell, L. A. Ehler, J. A. M. Mican, M. Baseler, et al., Presence of an inducible HIV-1 latent reservoir during highly active antiretroviral therapy, Proc. Natl. Acad. Sci. USA, 94 (1997), 13193-13197. |
| [36] | A. M. Elaiw, A. A. Raezah, S. A. Azoz, Stability of delayed HIV dynamics models with two latent reservoirs and immune impairment, Adv. Differ. Equ., 2018 (2018), Article Number: 414. |
| [37] | L. Huijuan, Z. Jia-Fang, Dynamics of two time delays differential equation model to HIV latent infection, Phys. A, 514 (2019), 384-395. |
| [38] | A. M. Elaiw, E. K. Elnahary, A. A. Raezah, Effect of cellular reservoirs and delays on the global dynamics of HIV, Adv. Differ. Equ., 2018 (2018), Article Number: 85. |
| [39] | B. Buonomo, C. Vargas-De-Leon, Global stability for an HIV-1 infection model including an eclipse stage of infected cells, J. Math. Anal. Appl., 385 (2012), 709-720. |
| [40] | A. M. Elaiw, M. A. Alshaikh, Stability of discrete-time HIV dynamics models with three categories of infected CD4+ T-cells, Adv. Differ. Equ., 2019 (2019), Article Number: 407. |
| [41] | L. Agosto, M. Herring, W. Mothes, A. Henderson, HIV-1-infected CD4+ T cells facilitate latent infection of resting CD4+ T cells through cell-cell contact, Cell, 24 (2018), 2088-2100. |
| [42] |
A. M. Elaiw, N. H. AlShamrani, Stability of a general adaptive immunity virus dynamics model with multi-stages of infected cells and two routes of infection, Math. Methods Appl. Sci., 43 (2020), 1145-1175. doi: 10.1002/mma.5923
|
| [43] |
X. Wang, S. Tang, X. Song, L. Rong, Mathematical analysis of an HIV latent infection model including both virus-to-cell infection and cell-to-cell transmission, J. Biol. Dyn., 11 (2017), 455-483. doi: 10.1080/17513758.2016.1242784
|
| [44] | A. D. Hobiny, A. M. Elaiw, A. Almatrafi, Stability of delayed pathogen dynamics models with latency and two routes of infection, Adv. Differ. Equ., 2018 (2018), Article Number: 276. |
| [45] | A. M. Elaiw, N. H. AlShamrani, Global stability of a delayed adaptive immunity viral infection with two routes of infection and multi-stages of infected cells, Commun. Nonlin. Sci. Numer. Simul., 86 (2020), Article ID 105259. |
| [46] |
W. Wang, X. Wang, K. Guo, W. Ma, Global analysis of a diffusive viral model with cell-to-cell infection and incubation period, Math. Methods Appl. Sci., 43 (2020), 5963-5978. doi: 10.1002/mma.6339
|
| [47] | K. Hattaf, H. Dutta, Modeling the dynamics of viral infections in presence of latently infected cells, Chaos Solitons Fractals, 136 (2020), Article ID 109916. |
| [48] | A. M. Elaiw, N. H. AlShamrani, Stability of a general CTL-mediated immunity HIV infection model with silent infected cell-to-cell spread, Adv. Differ. Equ., 2020 (2020), Article Number: 355. |
| [49] | J. K. Hale, S. V. Lunel, Introduction to functional differential equations, Springer-Verlag, New York, (1993). |
| [50] | Y. Kuang, Delay Differential Equations with Applications in Population Dynamics, San Diego: Academic Press, (1993). |
| [51] |
A. Korobeinikov, Global properties of infectious disease models with nonlinear incidence, Bull. Math. Biol., 69 (2007), 1871-1886. doi: 10.1007/s11538-007-9196-y
|
| [52] |
A. Korobeinikov, Global asymptotic properties of virus dynamics models with dose-dependent parasite reproduction and virulence and non-linear incidence rate, Math. Med. Biol., 26 (2009), 225-239. doi: 10.1093/imammb/dqp006
|
| [53] | X. Zhou, X. Shi, Z. Zhang, X. Song, Dynamical behavior of a virus dynamics model with CTL immune response, Appl. Math. Comput., 213(2009), 329-347. |
| [54] | X. Yang, L. S. Chen, J. F. Chen, Permanence and positive periodic solution for the single-species nonautonomous delay diffusive models, Comput. Math. Appl., 32 (1996), 109-116. |
| [55] |
P. van den Driessche, J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002), 29-48. doi: 10.1016/S0025-5564(02)00108-6
|
| [56] |
A. Korobeinikov, Global properties of basic virus dynamics models, Bull. Math. Biol., 66 (2004), 879-883. doi: 10.1016/j.bulm.2004.02.001
|
| [57] | A. M. Elaiw, S. F. Alshehaiween, A. D. Hobiny, Global properties of a delay-distributed HIV dynamics model including impairment of B-cell functions, Mathematics, 7 (2019) Article Number: 837. |
| [58] |
A. M. Elaiw, I. A. Hassanien, S. A. Azoz, Global stability of HIV infection models with intracellular delays, J. Korean Math. Soc., 49 (2012), 779-794. doi: 10.4134/JKMS.2012.49.4.779
|
| [59] | A. M. Elaiw, E. K. Elnahary, Analysis of general humoral immunity HIV dynamics model with HAART and distributed delays, Mathematics, 7 (2019), Article Number: 157. |
| [60] |
H. Shu, Y. Chen, L.Wang, Impacts of the cell-free and cell-to-cell infection modes on viral dynamics, J. Dyn. Differ. Equ., 30 (2018), 1817-1836. doi: 10.1007/s10884-017-9622-2
|
| [61] |
A. S. Perelson, P. W. Nelson, Mathematical analysis of HIV-1 dynamics in vivo, SIAM Rev., 41 (1999), 3-44. doi: 10.1137/S0036144598335107
|
| [62] |
P. D. Leenheer, H. L. Smith, Virus dynamics: A global analysis, SIAM J. Appl. Math., 63 (2003), 1313-1327. doi: 10.1137/S0036139902406905
|
| [63] | N. Bellomo, Y. Tao, Stabilization in a chemotaxis model for virus infection, Discrete Contin. Dyn. Syst. Ser. S, 13 (2020), 105-117. |
| [64] |
N. Bellomo, K. J. Painter, Y. Tao, M. Winkler, Occurrence vs. Absence of taxis-driven instabilities in a May-Nowak model for virus infection, SIAM J. Appl. Math., 79 (2019), 1990-2010. doi: 10.1137/19M1250261
|
| [65] |
C. Qin, Y. Chen, X. Wang, Global dynamics of a delayed diffusive virus infection model with cell-mediated immunity and cell-to-cell transmission, Math. Biosci. Eng., 17 (2020), 4678-4705. doi: 10.3934/mbe.2020257
|
| [66] | A. M. Elaiw, A. D. AlAgha, Global dynamics of reaction-diffusion oncolytic M1 virotherapy with immune response, Appl. Math. Comput., 367 (2020), Article 124758. |
| [67] | A. M. Elaiw, A. D. AlAgha, Global analysis of a reaction-diffusion within-host malaria infection model with adaptive immune response, Mathematics, 8 (2020), Article Number: 563. |
| [68] | A. M. Elaiw, A. D. AlAgha, Analysis of a delayed and diffusive oncolytic M1 virotherapy model with immune response, Nonlinear Anal. Real World Appl., 55 (2020), Article 103116. |
| [69] | L. Gibelli, A. Elaiw, M. A. Alghamdi, A. M. Althiabi, Heterogeneous population dynamics of active particles: Progression, mutations, and selection dynamics, Math. Models Methods Appl. Sci., 27 (2017), 617-640. |
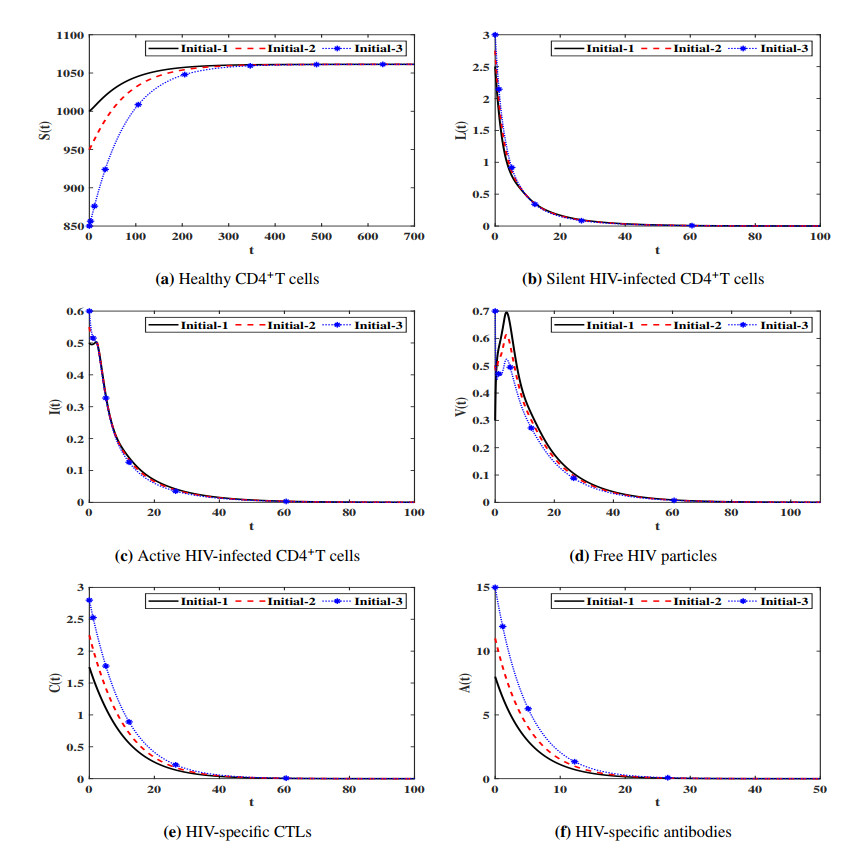
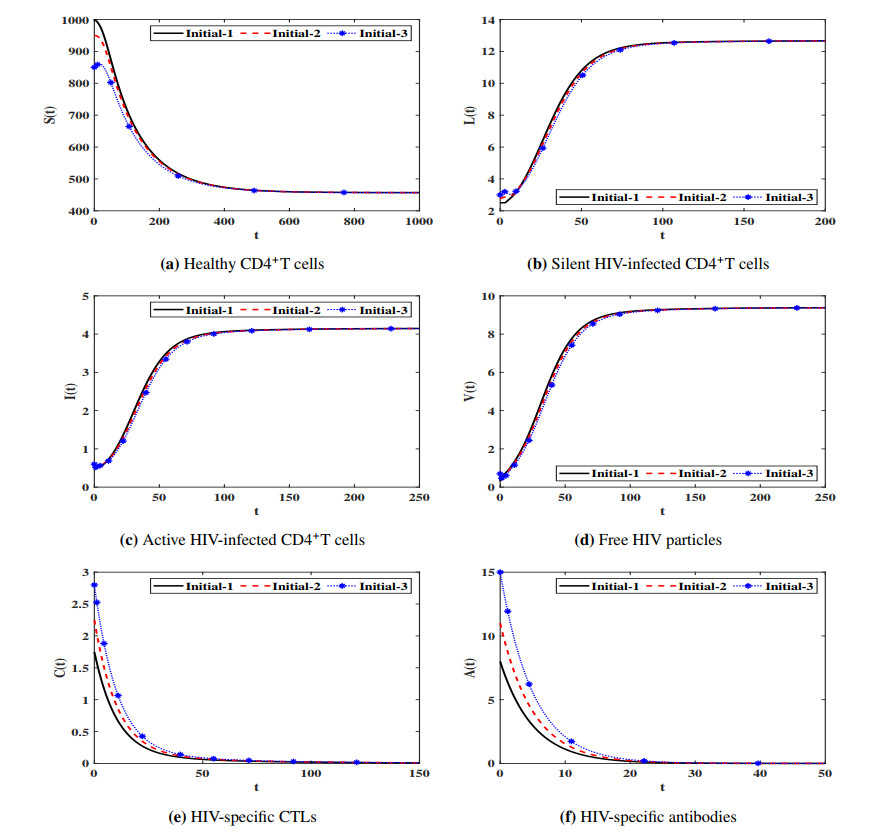
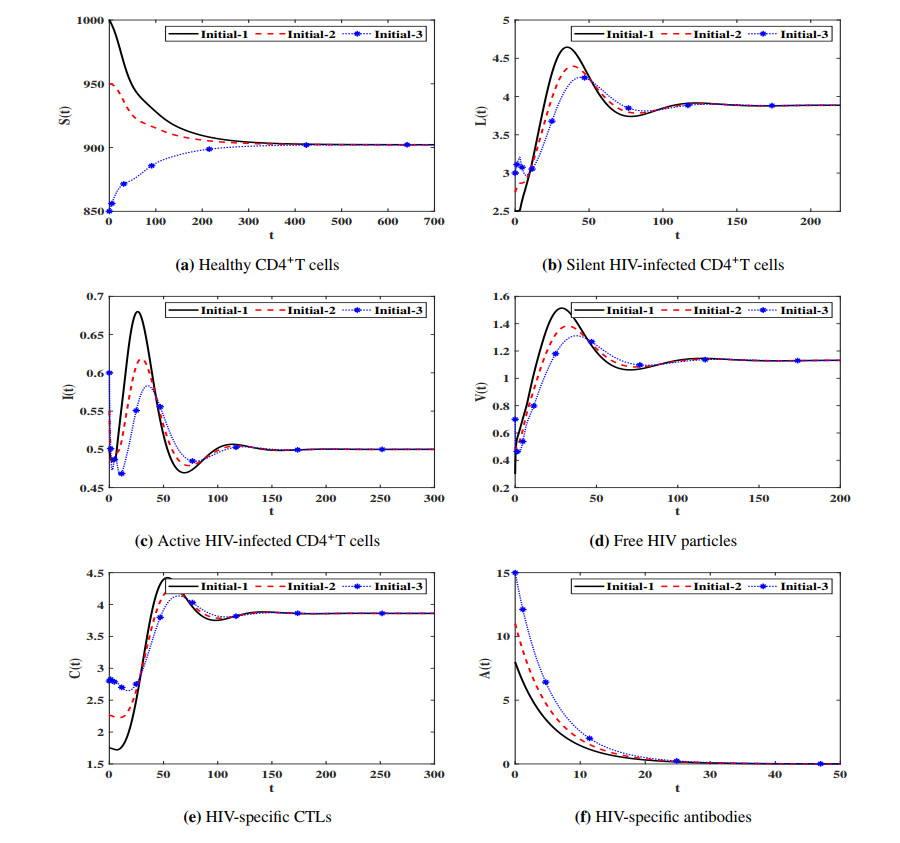
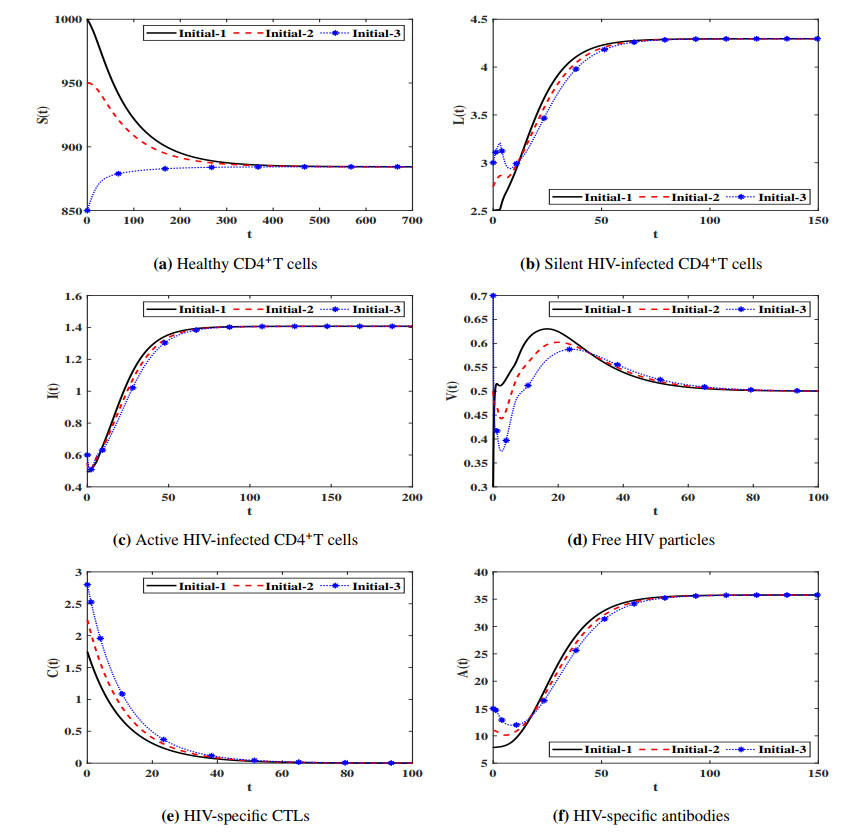
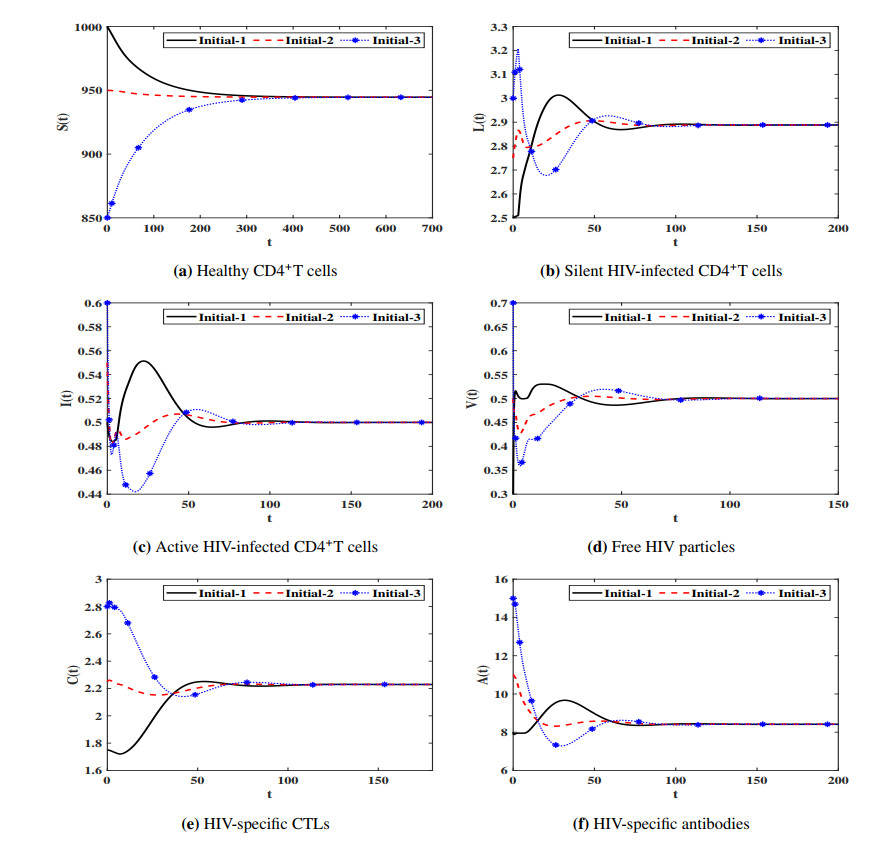
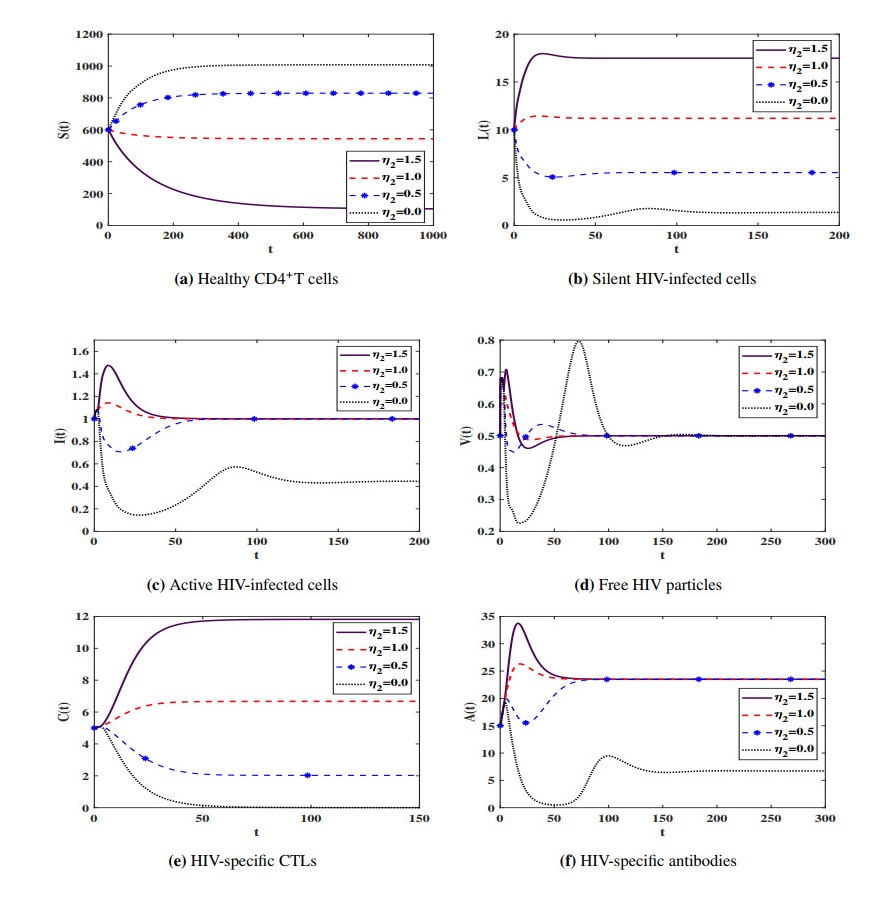
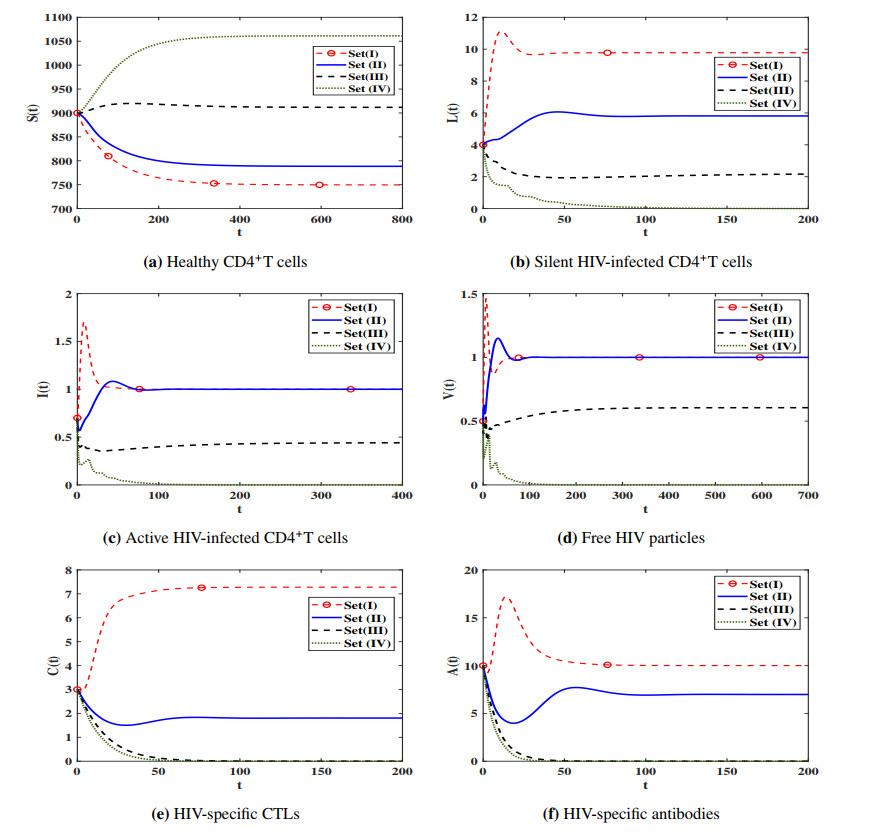
Figures(7) / Tables(4)

A. M. Elaiw, N. H. AlShamrani, A. D. Hobiny. Stability of an adaptive immunity delayed HIV infection model with active and silent cell-to-cell spread[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 6401-6458. doi: 10.3934/mbe.2020337







 DownLoad:
DownLoad: