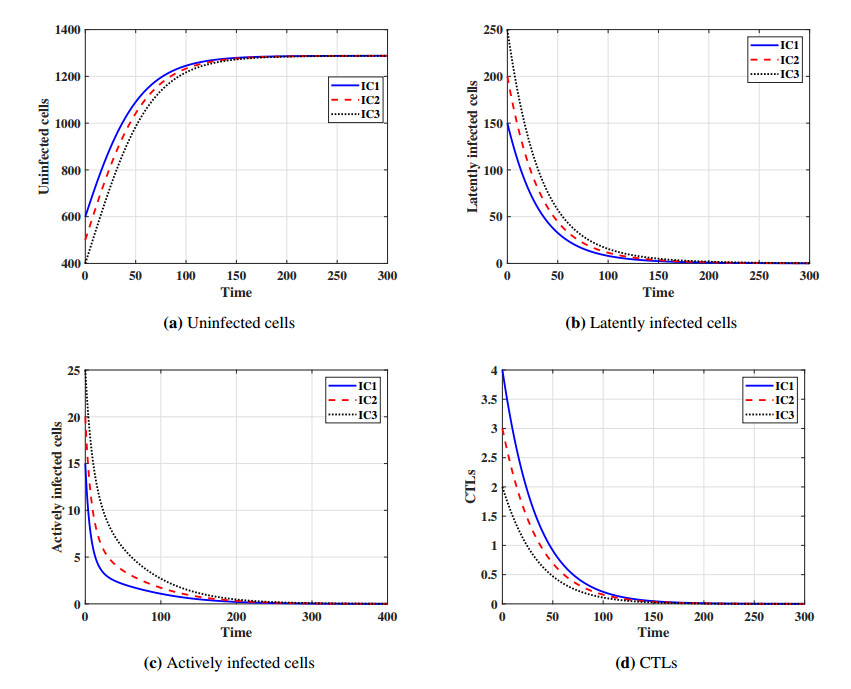
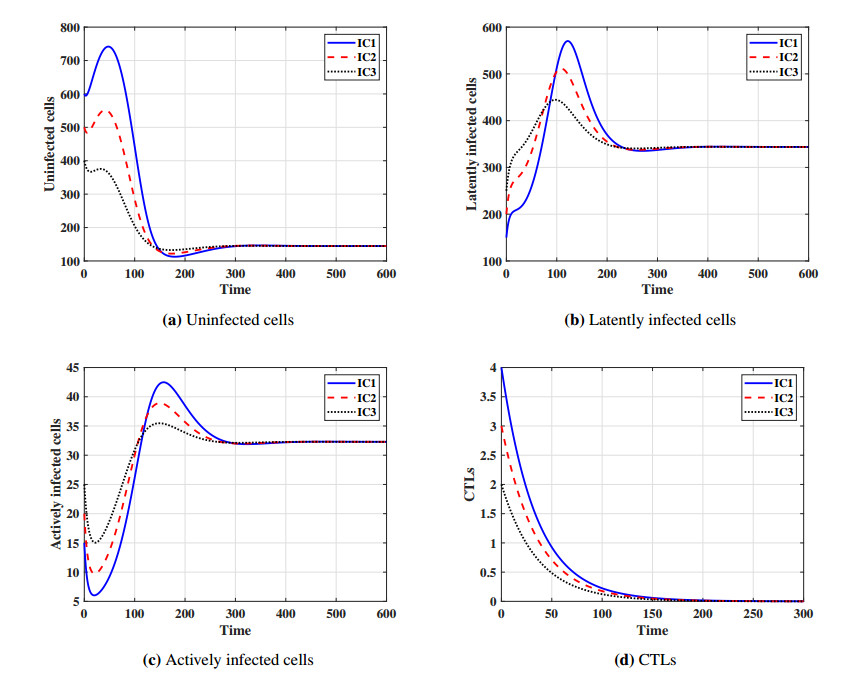
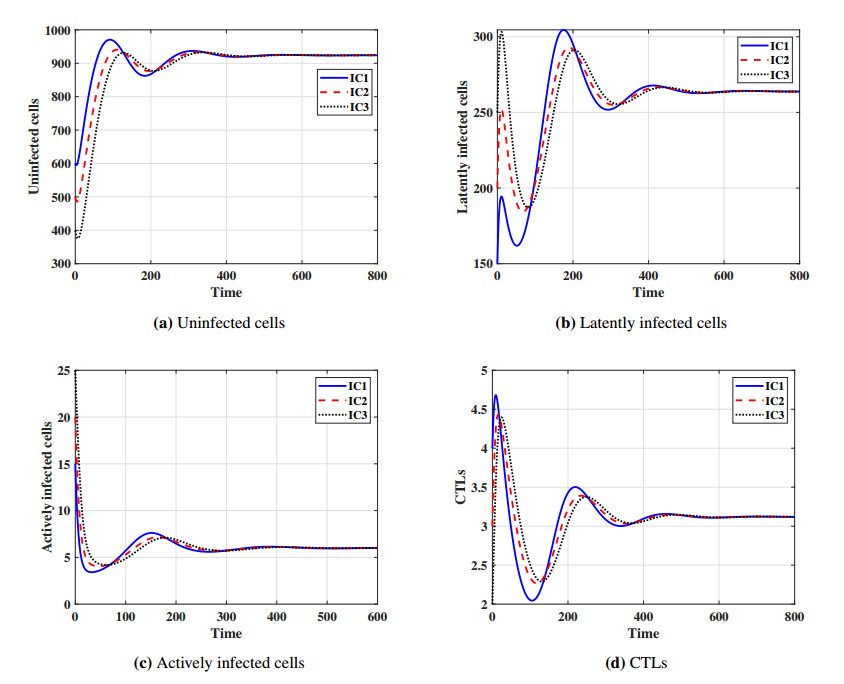
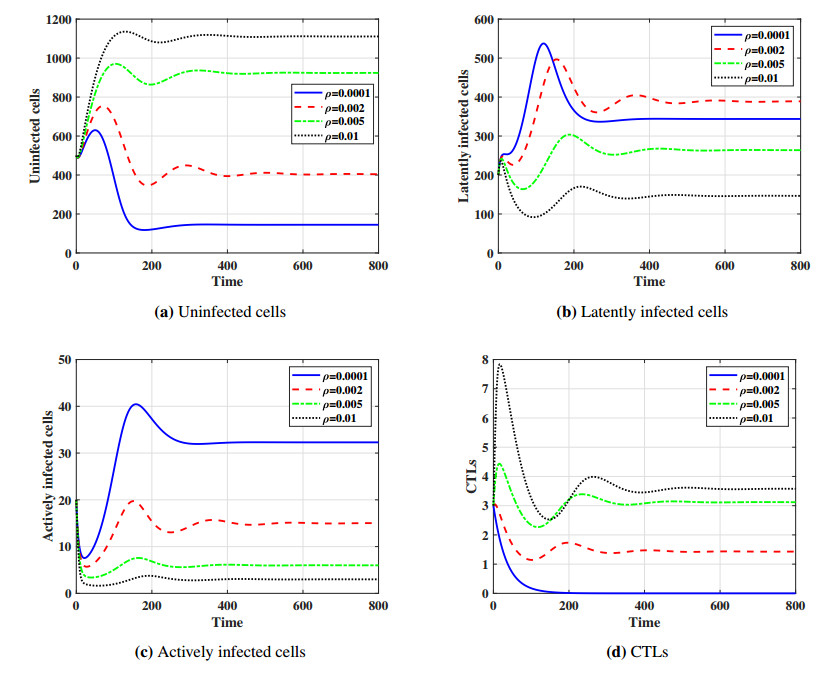
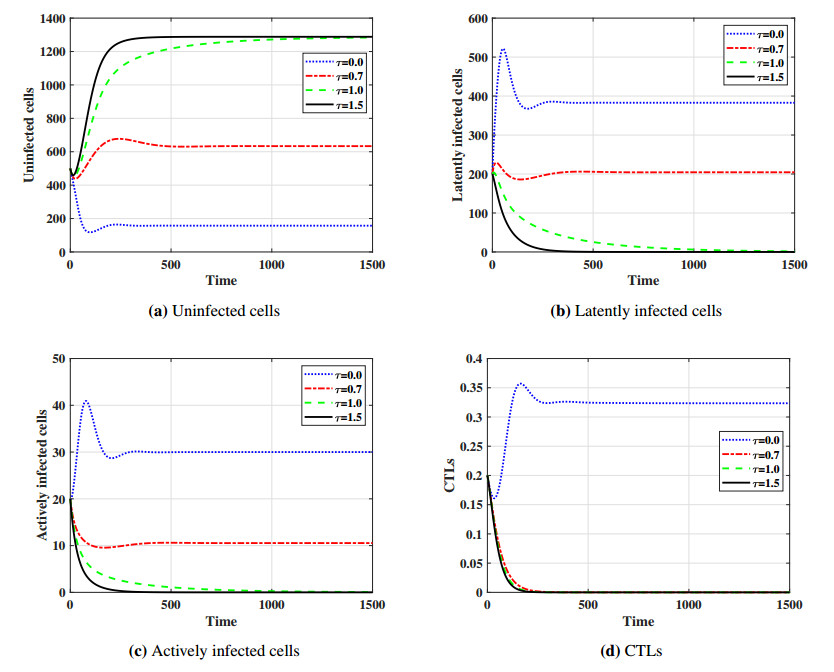
This paper formulates and analyzes a general delayed mathematical model which describe the within-host dynamics of Human T-cell lymphotropic virus class I (HTLV-I) under the effect Cytotoxic T Lymphocyte (CTL) immunity. The models consist of four components: uninfected CD$ 4^{+} $T cells, latently infected cells, actively infected cells and CTLs. The mitotic division of actively infected cells are modeled. We consider general nonlinear functions for the generation, proliferation and clearance rates for all types of cells. The incidence rate of infection is also modeled by a general nonlinear function. These general functions are assumed to be satisfy some suitable conditions. To account for series of events in the infection process and activation of latently infected cells, we introduce two intracellular distributed-time delays into the models: (ⅰ) delay in the formation of latently infected cells, (ⅱ) delay in the activation of latently infected cells. We determine a bounded domain for the system's solutions. We calculate two threshold numbers, the basic reproductive number $ R_{0} $ and the CTL immunity stimulation number $ R_{1} $. We determine the conditions for the existence and global stability of the equilibrium points. We study the global stability of all equilibrium points using Lyapunov method. We prove the following: (a) if $ R_{0}\leq 1 $, then the infection-free equilibrium point is globally asymptotically stable (GAS), (b) if $ R_{1}\leq 1 < R_{0} $, then the infected equilibrium point without CTL immunity is GAS, (c) if $ R_{1} > 1 $, then the infected equilibrium point with CTL immunity is GAS. We present numerical simulations for the system by choosing special shapes of the general functions. The effects of proliferation of CTLs and time delay on the HTLV-I progression is investigated. We noted that the CTL immunity does not play the role in clearing the HTLV-I from the body, but it has an important role in controlling and suppressing the viral infection. On the other hand, we observed that, increasing the time delay intervals can have similar influences as drug therapies in removing viruses from the body. This gives some impression to develop two types of treatments, the first type aims to extend the intracellular delay periods, while the second type aims to activate and stimulate the CTL immune response.
Citation: A. M. Elaiw, A. S. Shflot, A. D. Hobiny. Stability analysis of general delayed HTLV-I dynamics model with mitosis and CTL immunity[J]. Mathematical Biosciences and Engineering, 2022, 19(12): 12693-12729. doi: 10.3934/mbe.2022593
This paper formulates and analyzes a general delayed mathematical model which describe the within-host dynamics of Human T-cell lymphotropic virus class I (HTLV-I) under the effect Cytotoxic T Lymphocyte (CTL) immunity. The models consist of four components: uninfected CD$ 4^{+} $T cells, latently infected cells, actively infected cells and CTLs. The mitotic division of actively infected cells are modeled. We consider general nonlinear functions for the generation, proliferation and clearance rates for all types of cells. The incidence rate of infection is also modeled by a general nonlinear function. These general functions are assumed to be satisfy some suitable conditions. To account for series of events in the infection process and activation of latently infected cells, we introduce two intracellular distributed-time delays into the models: (ⅰ) delay in the formation of latently infected cells, (ⅱ) delay in the activation of latently infected cells. We determine a bounded domain for the system's solutions. We calculate two threshold numbers, the basic reproductive number $ R_{0} $ and the CTL immunity stimulation number $ R_{1} $. We determine the conditions for the existence and global stability of the equilibrium points. We study the global stability of all equilibrium points using Lyapunov method. We prove the following: (a) if $ R_{0}\leq 1 $, then the infection-free equilibrium point is globally asymptotically stable (GAS), (b) if $ R_{1}\leq 1 < R_{0} $, then the infected equilibrium point without CTL immunity is GAS, (c) if $ R_{1} > 1 $, then the infected equilibrium point with CTL immunity is GAS. We present numerical simulations for the system by choosing special shapes of the general functions. The effects of proliferation of CTLs and time delay on the HTLV-I progression is investigated. We noted that the CTL immunity does not play the role in clearing the HTLV-I from the body, but it has an important role in controlling and suppressing the viral infection. On the other hand, we observed that, increasing the time delay intervals can have similar influences as drug therapies in removing viruses from the body. This gives some impression to develop two types of treatments, the first type aims to extend the intracellular delay periods, while the second type aims to activate and stimulate the CTL immune response.

| [1] |
S. Khajanchi, S. Bera, T. K. Roy, Mathematical analysis of the global dynamics of a HTLV-I infection model, considering the role of cytotoxic T-lymphocytes, Math. Comput. Simul., 180 (2021), 354–378. https://doi.org/10.1016/j.matcom.2020.09.009 doi: 10.1016/j.matcom.2020.09.009

|
| [2] |
F. A. Proietti, A. B. F. Carneiro-Proietti, B. C. Catalan-Soares, E. L. Murphy, Global epidemiology of HTLV-I infection and associated diseases, Oncogene, 24 (2005), 6058–6068. https://doi.org/10.1038/sj.onc.1208968 doi: 10.1038/sj.onc.1208968
|
| [3] |
C. R. M. Bangham, HTLV-I infections, J. Clin. Pathol., 53 (2000), 581–586. http://dx.doi.org/10.1136/jcp.53.8.581 doi: 10.1136/jcp.53.8.581
|
| [4] |
D. Wodarz, C. R. M. Bangham, Evolutionary dynamics of HTLV-I, J. Mol. Evol., 50 (2000), 448–455. https://doi.org/10.1007/s002390010047 doi: 10.1007/s002390010047
|
| [5] |
B. Asquith, C. R. M. Bangham, How does HTLV-I persist despite a strong cell-mediated immune response?, Trends Immunol., 29 (2008), 4–11. https://doi.org/10.1016/j.it.2007.09.006 doi: 10.1016/j.it.2007.09.006
|
| [6] | H. Shiraki, Y. Sagara, Y. Inoue, Cell-to-cell transmission of HTLV-I, Gann Monogr. Cancer Res., 50 (2003), 303–316. |
| [7] |
N. I. Stilianakis, J. Seydel, Modeling the T-cell dynamics and pathogenesis of HTLV-I infection, Bull. Math. Biol., 61 (1999), 935–947. https://doi.org/10.1006/bulm.1999.0117 doi: 10.1006/bulm.1999.0117
|
| [8] |
H. Gomez-Acevedo, M. Y. Li, Backward bifurcation in a model for HTLV-I infection of CD4$^{+}$T cells, Bull. Math. Biol., 67 (2005), 101–114. https://doi.org/10.1016/j.bulm.2004.06.004 doi: 10.1016/j.bulm.2004.06.004
|
| [9] |
C. Vargas-De-Leon, The complete classification for global dynamics of a model for the persistence of HTLV-1 infection, Appl. Math. Comput., 237 (2014), 489–493. https://doi.org/10.1016/j.amc.2014.03.138 doi: 10.1016/j.amc.2014.03.138
|
| [10] |
M. Y. Li, A. G. Lim, Modelling the role of tax expression in HTLV-I persistence in vivo, Bull. Math. Biol., 73 (2011), 3008–3029. https://doi.org/10.1007/s11538-011-9657-1 doi: 10.1007/s11538-011-9657-1
|
| [11] |
X. Song, Y. Li, Global stability and periodic solution of a model for HTLV-1 infection and ATL progression, Appl. Math. Comput., 180 (2006), 401–410. https://doi.org/10.1016/j.amc.2005.12.022 doi: 10.1016/j.amc.2005.12.022
|
| [12] |
L. Wang, M. Y. Li, D. Kirschner, Mathematical analysis of the global dynamics of a model for HTLV-I infection and ATL progression, Math. Biosci., 179 (2002), 207–217. https://doi.org/10.1016/S0025-5564(02)00103-7 doi: 10.1016/S0025-5564(02)00103-7
|
| [13] | B. Asquith, C. R. M. Bangham, Quantifying HTLV-I dynamics, Immunol. Cell Biol., 85 (2007), 280–286. https://doi.org/10.1038/sj.icb.7100050 |
| [14] |
A. G. Lim, P. K. Maini, HTLV-I infection: A dynamic struggle between viral persistence and host immunity, J. Theor. Biol., 352 (2014), 92–108. https://doi.org/10.1016/j.jtbi.2014.02.022 doi: 10.1016/j.jtbi.2014.02.022
|
| [15] |
C. R. M. Bangham, CTL quality and the control of human retroviral infections, Eur. J. Immunol., 39 (2009), 1700–1712. https://doi.org/10.1002/eji.200939451 doi: 10.1002/eji.200939451
|
| [16] |
X. Pan, Y. Chen, H. Shu, Rich dynamics in a delayed HTLV-I infection model: Stability switch, multiple stable cycles, and torus, J. Math. Anal. Appl., 479 (2019), 2214–2235. https://doi.org/10.1016/j.jmaa.2019.07.051 doi: 10.1016/j.jmaa.2019.07.051
|
| [17] |
C. Bartholdy, J. P. Christensen, D. Wodarz, A. R. Thomsen, Persistent virus infection despite chronic cytotoxic T-lymphocyte activation in gamma interferon-deficient mice infected with lymphocytic choriomeningitis virus, J. Virol., 74 (2000), 10304–10311. https://doi.org/10.1128/JVI.74.22.10304-10311.2000 doi: 10.1128/JVI.74.22.10304-10311.2000
|
| [18] |
H. Gomez-Acevedo, M. Y. Li, S. Jacobson, Multi-stability in a model for CTL response to HTLV-I infection and its implications to HAM/TSP development and prevention, Bull. Math. Biol., 72 (2010), 681–696. https://doi.org/10.1007/s11538-009-9465-z doi: 10.1007/s11538-009-9465-z
|
| [19] |
J. Lang, M. Y. Li, Stable and transient periodic oscillations in a mathematical model for CTL response to HTLV-I infection, J. Math. Biol., 65 (2012), 181–199. https://doi.org/10.1007/s00285-011-0455-z doi: 10.1007/s00285-011-0455-z
|
| [20] |
M. Y. Li, H. Shu, Multiple stable periodic oscillations in a mathematical model of CTL response to HTLV-I infection, Bull. Math. Biol., 73 (2011), 1774–1793. https://doi.org/10.1007/s11538-010-9591-7 doi: 10.1007/s11538-010-9591-7
|
| [21] |
M. Y. Li, H. Shu, Global dynamics of a mathematical model for HTLV-I infection of CD4+ T cells with delayed CTL response, Nonlinear Anal. Real World Appl., 13 (2012), 1080–1092. https://doi.org/10.1016/j.nonrwa.2011.02.026 doi: 10.1016/j.nonrwa.2011.02.026
|
| [22] |
D. Wodarz, M. A. Nowak, C. R. M. Bangham, The dynamics of HTLV-I and the CTL response, Immunol. Today, 20 (1999), 220–227. https://doi.org/10.1016/S0167-5699(99)01446-2 doi: 10.1016/S0167-5699(99)01446-2
|
| [23] |
L. Wang, Z. Liu, Y. Li, D. Xu, Complete dynamical analysis for a nonlinear HTLV-I infection model with distributed delay, CTL response and immune impairment, Discrete Contin. Dyn. Syst., 25 (2020), 917–933. http://dx.doi.org/10.3934/dcdsb.2019196 doi: 10.3934/dcdsb.2019196
|
| [24] |
Y. Muroya, Y. Enatsu, H. Li, Global stability of a delayed HTLV-I infection model with a class of nonlinear incidence rates and CTLs immune response, Appl. Math. Comput., 219 (2013), 10559–10573. https://doi.org/10.1016/j.amc.2013.03.081 doi: 10.1016/j.amc.2013.03.081
|
| [25] |
Y. Wang, J. Liu, J. M. Heffernan, Viral dynamics of an HTLV-I infection model with intracellular delay and CTL immune response delay, J. Math. Anal. Appl., 459 (2018), 506–527. https://doi.org/10.1016/j.jmaa.2017.10.027 doi: 10.1016/j.jmaa.2017.10.027
|
| [26] |
F. Li, W. Ma, Dynamics analysis of an HTLV-1 infection model with mitotic division of actively infected cells and delayed CTL immune response, Math. Methods Appl. Sci., 41 (2018), 3000–3017. https://doi.org/10.1002/mma.4797 doi: 10.1002/mma.4797
|
| [27] |
S. Li, Y. Zhou, Backward bifurcation of an HTLV-I model with immune response, Discrete Contin. Dyn. Syst., 21 (2016), 863–881. http://dx.doi.org/10.3934/dcdsb.2016.21.863 doi: 10.3934/dcdsb.2016.21.863
|
| [28] |
W. Wang, W. Ma, Global dynamics of a reaction and diffusion model for an HTLV-I infection with mitotic division of actively infected cells, J. Appl. Anal. Comput., 7 (2017), 899–930. http://dx.doi.org/10.11948/2017057 doi: 10.11948/2017057
|
| [29] |
X. Jia, R. Xu, Global dynamics of a delayed HTLV-I infection model with Beddington-DeAngelis incidence and immune impairment, Chaos Solitons Fractals, 155 (2022), 111733. https://doi.org/10.1016/j.chaos.2021.111733 doi: 10.1016/j.chaos.2021.111733
|
| [30] |
A. M. Elaiw, N. H. AlShamrani, Stability of HIV/HTLV-I co-infection model with delays, Math. Methods Appl. Sci., 45 (2022), 238–300. https://doi.org/10.1002/mma.7775 doi: 10.1002/mma.7775
|
| [31] |
P. Katri, S. Ruan, Dynamics of human T-cell lymphotropic virus I (HTLV-I) infection of CD$4^{+}$T cells, C. R. Biol., 327 (2004), 1009–1016. https://doi.org/10.1016/j.crvi.2004.05.011 doi: 10.1016/j.crvi.2004.05.011
|
| [32] |
Y. Wang, J. Liu, Global stability for delay-dependent HTLV-I model with CTL immune response, AIP Conf. Proc., 1738 (2016), 480074. https://doi.org/10.1063/1.4952310 doi: 10.1063/1.4952310
|
| [33] | X. Sun, J. Wei, Global dynamics of a HTLV-I infection model with CTL response, Elec. J. Qual. Theory Diff. Equations, 2013 (2013), 1–15. |
| [34] | M. Y. Li, X. Lin, H. Wang, Global Hopf branches in a delayed model for immune response to HTLV-1 infections: coexistence of multiple limit cycles, Can. Appl. Math. Q., 20 (2012), 39–50. |
| [35] |
S. Bera, S. Khajanchi, T. K. Roy, Dynamics of an HTLV-I infection model with delayed CTLs immune response, Appl. Math. Comput., 430 (2022), 127206. https://doi.org/10.1016/j.amc.2022.127206 doi: 10.1016/j.amc.2022.127206
|
| [36] |
X. Lu, L. Hui, S. Liu, J. Li, A mathematical model of HTLV-I infection with two time delays, Math. Biosci. Eng., 12 (2015), 431–449. http://dx.doi.org/10.3934/mbe.2015.12.431 doi: 10.3934/mbe.2015.12.431
|
| [37] |
A. Korobeinikov, Global asymptotic properties of virus dynamics models with dose-dependent parasite reproduction and virulence and non-linear incidence rate, Math. Med. Biol. J. IMA, 26 (2009), 225–239. https://doi.org/10.1093/imammb/dqp006 doi: 10.1093/imammb/dqp006
|
| [38] |
K. Qi, D. Jiang, Threshold behavior in a stochastic HTLV-I infection model with CTL immune response and regime switching, Math. Methods Appl. Sci., 41 (2018), 6866–6882. https://doi.org/10.1002/mma.5198 doi: 10.1002/mma.5198
|
| [39] |
L. Cai, X. Li, M. Ghosh, Global dynamics of a mathematical model for HTLV-I infection of CD4$^{+}$ T-cells, Appl. Math. Modell., 35 (2011), 3587–3595. https://doi.org/10.1016/j.apm.2011.01.033 doi: 10.1016/j.apm.2011.01.033
|
| [40] | J. Hale, S. M. V. Lunel, Introduction to Functional Differential Equations, Applied Mathematical Science, Springer Verlag, New York, 1993. |
| [41] |
A. M. Elaiw, N. H. AlShamrani, Modeling and analysis of a within-host HIV/HTLV-I co-infection, Bol. Soc. Mat. Mex., 27 (2021), 27–38. https://doi.org/10.1007/s40590-021-00330-6 doi: 10.1007/s40590-021-00330-6
|
| [42] |
A. M. Elaiw, N. H. AlShamrani, Analysis of a within-host HIV/HTLV-I co-infection model with immunity, Virus Res., 295 (2021), 198204. https://doi.org/10.1016/j.virusres.2020.198204 doi: 10.1016/j.virusres.2020.198204
|
| [43] |
X. Yang, L. Chen, J. Chen, Permanence and positive periodic solution for the single-species nonautonomous delay diffusive models, Comput. Math. Appl., 32 (1996), 109–116. https://doi.org/10.1016/0898-1221(96)00129-0 doi: 10.1016/0898-1221(96)00129-0
|
| [44] |
A. Korobeinikov, Global properties of basic virus dynamics models, Bull. Math. Biol., 66 (2004), 879–883. https://doi.org/10.1016/j.bulm.2004.02.001 doi: 10.1016/j.bulm.2004.02.001
|
| [45] |
A. Korobeinikov, Global properties of infectious disease models with nonlinear incidence, Bull. Math. Biol., 69 (2007), 1871–1886. https://doi.org/10.1007/s11538-007-9196-y doi: 10.1007/s11538-007-9196-y
|
| [46] |
A. M. Elaiw, N. H. AlShamrani, Global stability of humoral immunity virus dynamics models with nonlinear infection rate and removal, Nonlinear Anal. Real World Appl., 26 (2015), 161–190. https://doi.org/10.1016/j.nonrwa.2015.05.007 doi: 10.1016/j.nonrwa.2015.05.007
|
| [47] | E. A. Barbashin, Introduction to the theory of stability, Wolters-Noordhoff, Groningen, 1970. |
| [48] | J. P. LaSalle, The Stability of Dynamical Systems, SIAM, Philadelphia, 1976. |
| [49] |
A. M. Lyapunov, The general problem of the stability of motion, Int. J. Control, 55 (1992), 531–534. https://doi.org/10.1080/00207179208934253 doi: 10.1080/00207179208934253
|
| [50] |
A. S. Perelson, D. E. Kirschner, R. de boer, Dynamics of HIV Infection of CD4+ T cells, Math. Biosci., 114 (1993), 81–125. https://doi.org/10.1016/0025-5564(93)90043-A doi: 10.1016/0025-5564(93)90043-A
|
| [51] |
R. V. Culshaw, S. Ruan, A delay-differential equation model of HIV infection of CD4$^{+}$ T-cells, Math. Biosci., 165 (2000), 27–39. https://doi.org/10.1016/S0025-5564(00)00006-7 doi: 10.1016/S0025-5564(00)00006-7
|
| [52] |
Y. Wang, Y. Zhou, J. Wu, J. Heffernan, Oscillatory viral dynamics in a delayed HIV pathogenesis model, Math. Biosci., 219 (2009), 104–112. https://doi.org/10.1016/j.mbs.2009.03.003 doi: 10.1016/j.mbs.2009.03.003
|
| [53] |
M. N. Jan, N. Ali, G. Zaman, I. Ahmad, Z. Shah, P. Kumam, HIV-1 infection dynamics and optimal control with Crowley-Martin function response, Comput. Methods Prog. Biomed., 193 (2020), 105503. https://doi.org/10.1016/j.cmpb.2020.105503 doi: 10.1016/j.cmpb.2020.105503
|
| [54] |
B. Asquith, A. J. Mosley, A. Barfield, S. E. F. Marshall, A. Heaps, P. Goon, et al., A functional CD8$^{+}$ cell assay reveals individual variation in CD8 $^{+}$ cell antiviral efficacy and explains differences in human T-lymphotropic virus type 1 proviral load, J. Gener. Virol., 86 (2005), 1515–1523. https://doi.org/10.1099/vir.0.80766-0 doi: 10.1099/vir.0.80766-0
|
| [55] |
N. Bellomo, N. Outada, J. Soler, Y. Tao, M. Winkler, Chemotaxis and cross diffusion models in complex environments: Models and analytic problems toward a multiscale vision, Math. Models Methods Appl. Sci., 32 (2022), 713–792. https://doi.org/10.1142/S0218202522500166 doi: 10.1142/S0218202522500166
|
| [56] |
L. Gibelli, A. M. Elaiw, M. A. Alghamdi, A. M. Althiabi, Heterogeneous population dynamics of active particles: Progression, mutations, and selection dynamics, Math. Models Methods Appl. Sci., 27 (2017), 617–640. https://doi.org/10.1142/S0218202517500117 doi: 10.1142/S0218202517500117
|
Figures(5) / Tables(3)

A. M. Elaiw, A. S. Shflot, A. D. Hobiny. Stability analysis of general delayed HTLV-I dynamics model with mitosis and CTL immunity[J]. Mathematical Biosciences and Engineering, 2022, 19(12): 12693-12729. doi: 10.3934/mbe.2022593







 DownLoad:
DownLoad: