Citation: Pierre Degond, Maxime Herda, Sepideh Mirrahimi. A Fokker-Planck approach to the study of robustness in gene expression[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 6459-6486. doi: 10.3934/mbe.2020338

| [1] |
L. Bleris, Z. Xie, D. Glass, A. Adadey, E. Sontag, Y. Benenson, Synthetic incoherent feedforward circuits show adaptation to the amount of their genetic template, Molecular systems biology, 7 (2011), 519. doi: 10.1038/msb.2011.49

|
| [2] | R. Blevins, L. Bruno, T. Carroll, J. Elliott, A. Marcais, C. Loh, et al., Micrornas regulate cellto-cell variability of endogenous target gene expression in developing mouse thymocytes, PLoS genetics, 11 (2015). |
| [3] |
M. S. Ebert, P. A. Sharp, Roles for micrornas in conferring robustness to biological processes, Cell, 149 (2012), 515-524. doi: 10.1016/j.cell.2012.04.005
|
| [4] | H. Herranz, S. M. Cohen, Micrornas and gene regulatory networks: managing the impact of noise in biological systems, Genes & development, 24 (2010), 1339-1344. |
| [5] | M. Osella, C. Bosia, D. Corá, M. Caselle, The role of incoherent microrna-mediated feedforward loops in noise buffering, PLoS Comput Biol, 7 (2011), e1001101. |
| [6] | N. G. van Kampen, Stochastic Processes in Physics and Chemistry, North-Holland, Amsterdam, 1981. |
| [7] |
D. T. Gillespie, A general method for numerically simulating the stochastic time evolution of coupled chemical reactions, J. Comput. Phys., 22 (1976), 403-434. doi: 10.1016/0021-9991(76)90041-3
|
| [8] |
C. Bosia, M. Osella, M. El Baroudi, D. Corà, M. Caselle, Gene autoregulation via intronic micrornas and its functions, BMC Syst. Biol., 6 (2012), 131. doi: 10.1186/1752-0509-6-131
|
| [9] | P. Degond, S. Jin, Y. Zhu, An uncertainty quantification approach to the study of gene expression robustness, preprint, arXiv: 1910.07188. |
| [10] | B. Perthame, Parabolic Equations in Biology, Lecture Notes on Mathematical Modelling in the Life Sciences, Springer, Cham, 2015. |
| [11] | P. Lötstedt, L. Ferm, Dimensional reduction of the Fokker-Planck equation for stochastic chemical reactions, Multiscale Modeling & Simulation, 5 (2006), 593-614. |
| [12] | P. Degond, M. Herda, S. Mirrahimi, FPmuRNA, 2020. Available from: https://gitlab.inria.fr/herda/fpmurna. |
| [13] |
D. T. Gillespie, The chemical langevin equation, The Journal of Chemical Physics, 113 (2000), 297-306. doi: 10.1063/1.481811
|
| [14] |
W. Huang, M. Ji, Z. Liu, Y. Yi, Steady states of Fokker-Planck equations: I. Existence, J. Dynam. Differential Equations, 27 (2015), 721-742. doi: 10.1007/s10884-015-9454-x
|
| [15] |
W. Huang, M. Ji, Z. Liu, Y. Yi, Integral identity and measure estimates for stationary FokkerPlanck equations, Ann. Probab., 43 (2015), 1712-1730. doi: 10.1214/14-AOP917
|
| [16] | V. I. Bogachev, N. V. Krylov, M. Röckner, S. V. Shaposhnikov, Fokker-Planck-Kolmogorov Equations, American Mathematical Society, Providence, RI, 2015. |
| [17] | R. Z. Has'minskiĭ, Ergodic properties of recurrent diffusion processes and stabilization of the solution of the Cauchy problem for parabolic equations, Teor. Verojatnost. i Primenen., 5 (1960), 196-214. |
| [18] | D. Gilbarg, N. S. Trudinger, Elliptic Partial Differential Equations of Second Order, SpringerVerlag, Berlin, 2001. |
| [19] | F. W. J. Olver, D. W. Lozier, R. F. Boisvert, C. W. Clark, NIST Handbook of Mathematical Functions Hardback and CD-ROM, Cambridge University Press, Cambridge, 2010. |
| [20] | M. Bessemoulin-Chatard, M. Herda, T. Rey, Hypocoercivity and diffusion limit of a finite volume scheme for linear kinetic equations, Math. Comp., 89 (2020), 1093-1133. |
| [21] |
J. S. Chang, G. Cooper, A practical difference scheme for Fokker-Planck equations, Journal of Computational Physics, 6 (1970), 1-16. doi: 10.1016/0021-9991(70)90001-X
|
| [22] |
C. Chainais-Hillairet, J. Droniou, Finite-volume schemes for noncoercive elliptic problems with Neumann boundary conditions, IMA J. Numer. Anal., 31 (2011), 61-85. doi: 10.1093/imanum/drp009
|
| [23] | H. J. Brascamp, E. H. Lieb, On extensions of the Brunn-Minkowski and Prékopa-Leindler theorems, including inequalities for log concave functions, and with an application to the diffusion equation, J. Functional Analysis, 22 (1976), 366-389. |
| [24] |
S. G. Bobkov, M. Ledoux, From Brunn-Minkowski to Brascamp-Lieb and to logarithmic Sobolev inequalities, Geom. Funct. Anal., 10 (2000), 1028-1052. doi: 10.1007/PL00001645
|
| [25] | D. Bakry, Michel Émery, Diffusions hypercontractives, in Séminaire de Probabilités, XIX, 1983/84, Springer, Berlin, (1985), 177-206. |
| [26] | D. Bakry, L'hypercontractivité et son utilisation en théorie des semigroupes, in Lectures on Probability Theory (Saint-Flour, 1992), Springer, Berlin, (1994), 1-114. |
| [27] |
A. Arnold, P. Markowich, G. Toscani, A. Unterreiter, On convex Sobolev inequalities and the rate of convergence to equilibrium for Fokker-Planck type equations, Comm. Partial Differ. Equations, 26 (2001), 43-100. doi: 10.1081/PDE-100002246
|
| [28] |
A. Arnold, J. Dolbeault, Refined convex Sobolev inequalities, J. Funct. Anal., 225 (2005), 337-351. doi: 10.1016/j.jfa.2005.05.003
|
| [29] | B. Arras, Y. Swan, A stroll along the gamma, Stochastic Process. Appl., 127 (2017), 3661-3688. |
| [30] | D. Bakry, Remarques sur les semigroupes de Jacobi, Astérisque, 236 (1996), 23-39. |
| [31] |
M. Benaïm, R. Rossignol, Exponential concentration for first passage percolation through modified Poincaré inequalities, Ann. Inst. Henri Poincaré Probab. Stat., 44 (2008), 544-573. doi: 10.1214/07-AIHP124
|
| [32] | L. Miclo, Sur l'inégalité de Sobolev logarithmique des opérateurs de Laguerre à petit paramètre, in Séminaire de Probabilités, XXXVI, Springer, Berlin, (2003), 222-229. |
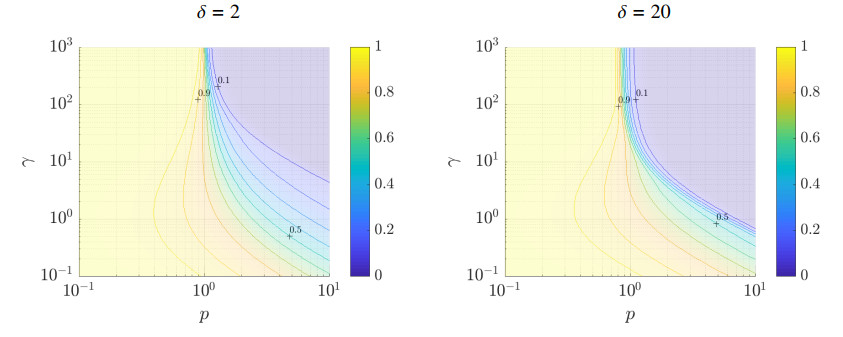
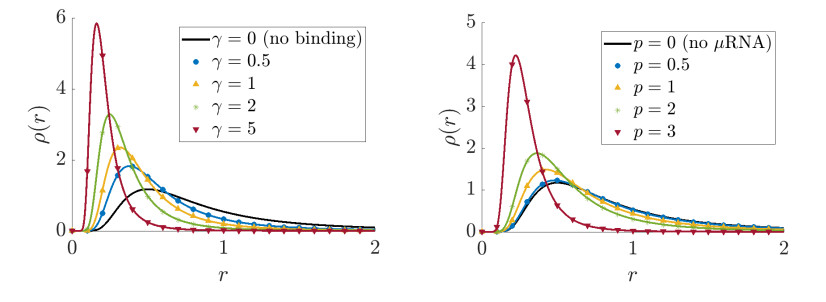
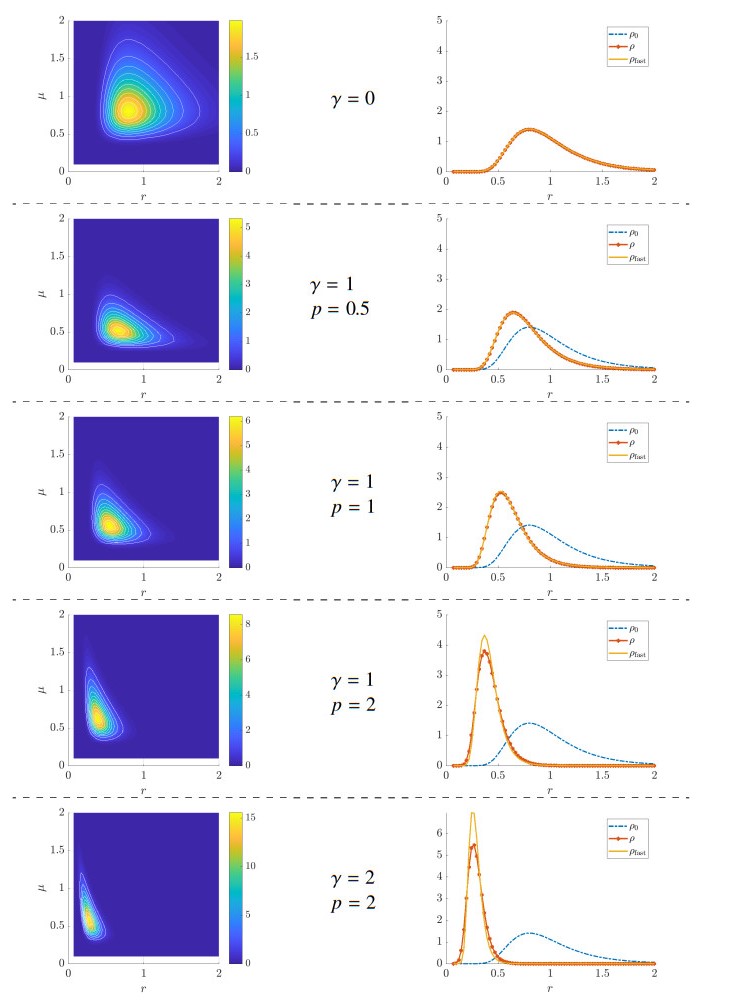
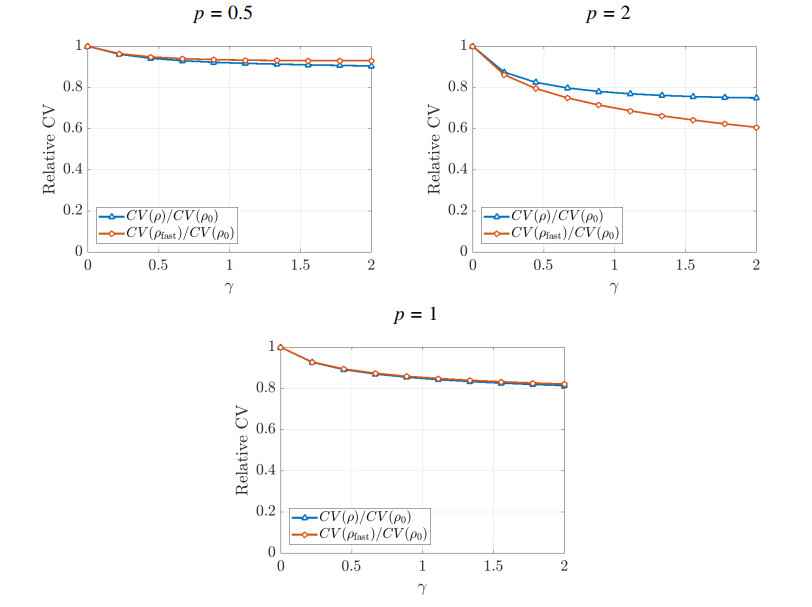
Figures(5)

Pierre Degond, Maxime Herda, Sepideh Mirrahimi. A Fokker-Planck approach to the study of robustness in gene expression[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 6459-6486. doi: 10.3934/mbe.2020338







 DownLoad:
DownLoad: