Citation: Huan Kong, Guohong Zhang, Kaifa Wang. Stability and Hopf bifurcation in a virus model with self-proliferation and delayed activation of immune cells[J]. Mathematical Biosciences and Engineering, 2020, 17(5): 4384-4405. doi: 10.3934/mbe.2020242

| [1] | J. E. Schmitz, M. J. Kuroda, S. Santra, V. G. Sasseville, M. A. Simon, M. A. Lifton, et al, Control of viremia in simian immunodeficiency virus infection by CD8+ lymphocytes, Science, 283 (1999), 857-860. |
| [2] | L. C. Wang, M. Y. Li, Mathematical analysis of the global dynamics of a model for HIV infection of CD4+ T cells, Math. Biosci., 200 (2006), 44-57. |
| [3] | X. Y. Song, A. U. Neumann, Global stability and periodic solution of the viral dynamics, J. Math. Anal. Appl., 329 (2007), 281-297. |
| [4] | R. J. De Boer, A. S. Perelson, Target cell limited and immune control models of HIV infection: A comparison, J. Theoret. Biol., 190 (1998), 201-214. |
| [5] | Y. Nakata, Global dynamics of a cell mediated immunity in viral infection models with distributed delays, J. Math. Anal. Appl., 375 (2011), 14-27. |
| [6] | J. L. Wang, J. M. Pang, T. Kuniya, Global threshold dynamics in a five-dimensional virus model with cell-mediated, humoral immune responses and distributed delays, Appl. Math. Comput., 241 (2014), 298-316. |
| [7] | M. A. Nowak, S. Bonhoefier, A. M. Hill, R. Boehme, H. C. Thomas, H. McDade, Viral dynamics in hepatitis B virus infection, Proc. Natl. Acad. Sci. USA, 93 (1996), 4398-4402. |
| [8] | A. Korobeinikov, S. Giles, Global properties of basic virus dynamics models, Bull. Math. Biol., 66 (2004), 879-883. |
| [9] | M. A. Nowak, C. R. M. Bangham, Population dynamics of immune responses to persistent viruses, Science, 272 (1996), 74-79. |
| [10] | H. Zhu, X. Zou, Dynamics of a HIV-1 Infection model with cell-mediated immune response and intracellular delay,Discr. Cont. Dyn. Syst. Ser. B, 12 (2009), 511-524. |
| [11] | K. Wang, W. Wang, H. Pang, X. Liu, Complex dynamic behavior in a viral model with delayed immune response, Physica D, 226 (2007), 197-208. |
| [12] | Yukihiko Nakata, Global dynamics of a cell mediated immunity in viral infection models with distributed delays, J. Math. Anal. Appl., 375 (2011), 14-27. |
| [13] | H. Gomez-Acevedo, M. Y. Li, S. Jacobson, Multi-stability in a model for CTL response to HTLVI infection and its consequences in HAM/TSP development and prevention, Bull. Math. Biol., 72 (2010), 681-696. |
| [14] | R. M. Anderson, R. M. May, S. Gupta, Non-linear phenomena in host-parasite interactions, Parasitology, 99 (1989), 59-79. |
| [15] | A. Murase, T. Sasaki, T. Kajiwara, Stability analysis of pathogen-immune interaction dynamics, J. Math. Biol., 51 (2005), 247-267. |
| [16] | C. Chiyaka, W. Garira, S. Dube, Modelling immune response and drug therapy in human malaria infection, Comput. Math. Method. Med., 9 (2008), 143-163. |
| [17] | A. S. Perelson, Modelling viral and immune system dynamics, Nature Rev. Immunol., 2 (2002), 28-36. |
| [18] | A. Korobeinikov, Immune response and within-host viral evolution:Immune response can accelerate evolution, J. Theor. Biol., 456 (2018),74-83. |
| [19] | H. Q. Zhang, H. Chen, C. C Jiang, K. F. Wang, Effect of explicit dynamics of free virus and intracellular delay, Chaos, Solitons Fractals, 104 (2017), 827-834. |
| [20] | Y. Wang, J. Liu, J. M. Heffernan, Viral dynamics of an HTLV-I infection model with intracellular delay and CTL immune response delay, J. Math. Anal. Appl., 459 (2018), 506-527. |
| [21] | K. Allali, S. Harroudi, D. F. M. Torre, Analysis and optimal control of an intracellular delayed HIV model with CTL immune response, Math. Comput. Sci., 12 (2018), 111-127. |
| [22] | H. J. Liu, J. F. Zhang, Dynamics of two time delays differential equation model to HIV latent infection, Physica A, 514 (2019), 384-395. |
| [23] | M. Y. Li, H. Shu, Multiple stable periodic oscillations in a mathematical model of CTL response to HTLV-I infection, Bull. Math. Biol., 73 (2011), 1774-1793. |
| [24] | D. W. Huang, X. Zhang, Y. F. Guo, H. L. Wang, Analysis of an HIV infection model with treatment sand delayed immune response, Appl. Math. Model., 40 (2016), 3081-3089. |
| [25] | D. Wodarz, J. P. Christensen, A. R. Thomsen, The importance of lytic and nonlytic immune responses in viral infections, Trends Immunol., 23 (2002), 194-200. |
| [26] | C. Bartholdy, J. P. Christensen, D. Wodarz, A. R. Thomsen, Persistent virus infection despite chronic cytotoxic T-lymphocyte activation in Gamma interferon-deficient mice infected with lymphocytic chroriomeningitis virus, J. Virology, 74 (2000), 10304-10311. |
| [27] | K. Wang, Y. Kuang, Fluctuation and extinction dynamics in host-microparasite systems, Comm. Pure Appl. Anal., 10 (2011), 1537-1548. |
| [28] | S. Bonhoeffer, J. M. Coffin, M. A. Nowak, Human immunodeficiency virus drug therapy and virus load, J. Virology, 71 (1997), 3275-3278. |
| [29] | M. Nagumo, Uber die lage der integralkurven gewohnlicher differentialgleichungen, Proc. Phys. Math. Soc., 24 (1942), 551-559. |
| [30] | P. van den Driessche, J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002), 29-48. |
| [31] | B. Hassard, D. Kazarinoff, Y. Wan, Theory and Applications of Hopf Bifurcation, Cambridge: Cambridge University Press, 1981. |
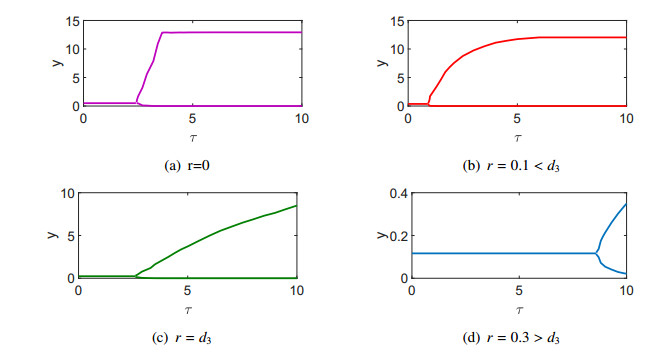
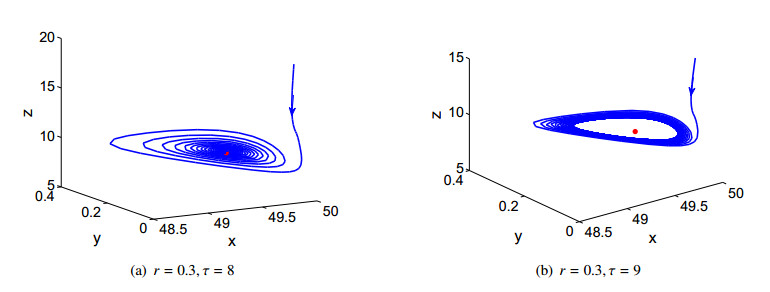
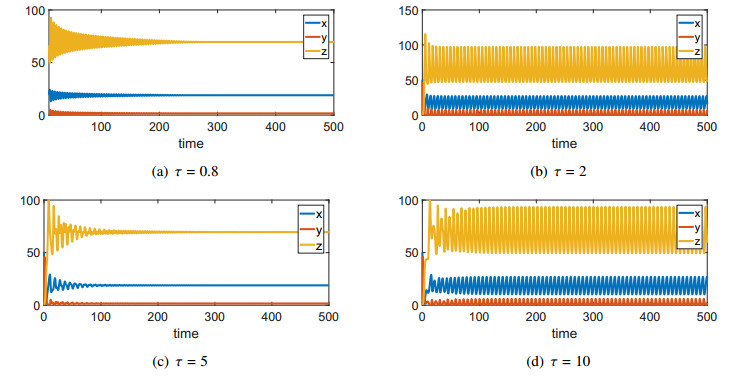
Figures(3) / Tables(2)

Huan Kong, Guohong Zhang, Kaifa Wang. Stability and Hopf bifurcation in a virus model with self-proliferation and delayed activation of immune cells[J]. Mathematical Biosciences and Engineering, 2020, 17(5): 4384-4405. doi: 10.3934/mbe.2020242







 DownLoad:
DownLoad: