Citation: Md Nazmul Hassan, Angela Peace. Mechanistically derived Toxicant-mediated predator-prey model under Stoichiometric constraints[J]. Mathematical Biosciences and Engineering, 2020, 17(1): 349-365. doi: 10.3934/mbe.2020019

| [1] | W. X. Wang and N. S. Fisher, Assimilation of trace elements and carbon by the mussel mytilus edulis:effects of food composition, Limnol. Oceanogr., 41 (1996), 197-207. |
| [2] | R. Ashauer and C. D. Brown, Toxicodynamic assumptions in ecotoxicological hazard models, Environ. Toxicol. Chem., 27 (2008), 1817-1821. |
| [3] | Q. Huang, L. Parshotam, H. Wang, et al., A model for the impact ofcontaminants on fish population dynamics. J. Theor. Biol., 334 (2013), 71-79. |
| [4] | Q. Huang, H. Wang and M. Lewis, Development of a toxin-mediated predator-prey model applicable to aquatic environments in the athabasca oil sands region, Osrin Report no. Tech. rep., (2014), TR-59. 59 pp. Available from: http://hdl.handle.net/10402/era.40140. |
| [5] | M. Danger and F. Maunoury-Danger, Ecological stoichiometry. In: Encyclopedia of Aquatic Ecotoxicology, Springer, (2013), 317-326. |
| [6] | O. Ieromina, W. J. Peijnenburg, G. de Snoo, et al., Impact of imidacloprid on daphnia magna under different food quality regimes, Environ. Toxicol. Chem., 33 (2014), 621-631. |
| [7] | C. R. Lessard and P. C. Frost, Phosphorus nutrition alters herbicide toxicity on Daphnia magna, Sci. Total Environ., 421 (2012), 124-128. |
| [8] | R. Karimi, C. Chen, P. Pickhardt, et al., Stoichiometric controls of mercury dilution by growth, P. Natl. Acad. Sci. USA, 104 (2007), 7477-7482. |
| [9] | A. Peace, M. Poteat and H. Wang, Somatic growth dilution of a toxicant in a predator-prey model under stoichiometric constraints, J. Theo. Biol., 407 (2016), 198-211. |
| [10] | M. N. Hassan, K. Thompson, G. Mayer, et al., Effect of excess food nutrient on producer-grazer model under stoichiometric and toxicological constraints, Math. Biosci. Eng., 16 (2018), 150-167. |
| [11] | R. W. Sterner and J. J. Elser, Ecological stoichiometry: the biology of elements from molecules to the biosphere, Princeton University Press, (2002). |
| [12] | A. Peace, Y. Zhao, I. Loladze, et al., A stoichiometric producer-grazer model incorporating the effects of excess food-nutrient content on consumer dynamics, Math. Biosci., 244 (2013), 107- 115. |
| [13] | F. J. Miller, P. M. Schlosser, D. B. Janszen, Haber's rule: a special case in a family of curves relating concentration and duration of exposure to a fixed level of response for a given endpoint, Toxicology, 149 (2000), 21-34. |
| [14] | T. Andersen, Pelagic nutrient cycles: herbivores as sources and sinks, Springer Science & Business Media, 129, 2013. |
| [15] | R. W. Vocke, K. L. Sears, J. J. O'Toole, et al., Growth responses of selected freshwater algae to trace elements and scrubber ash slurry generated by coal-fired power plants, Water Res., 14 (1980), 141-150. |
| [16] | K. E. Biesinger, L. E. Anderson and J. G. Eaton, Chronic effects of inorganic and organic mercury ondaphnia magna: Toxicity, accumulation, and loss, Arch. Environ. Con. Tox., 11 (1982), 769-774. |
| [17] | W. R. Hill and I. L. Larsen, Growth dilution of metals in microalgal biofilms, Environ. Sci. Technol., 39 (2005), 1513-1518. |
| [18] | M. T. Tsui and W. X. Wang, Uptake and elimination routes of inorganic mercury and methylmercury in daphnia magna, Environ. Sci. Technol., 38 (2004), 808-816. |
| [19] | B. Kooi, D. Bontje, G. Van Voorn, et al., Sublethal toxic effects in a simple aquatic foodchain, Ecol. Model., 212 (2008), 304-318. |
| [20] | I. Loladze, Y. Kuang and J. J. Elser, Stoichiometry in producer-grazer systems: Linking energy flow with element cycling, Bull. Math. Bio., 62L (2000), 1137-1162. |
| [21] | X. Li, H. Wang and Y. Kuang, Global analysis of a stoichiometric producer-grazer model withholling type functional responses, J. Math. Biol., 63 (2011), 901-932. |
| [22] | T. Xie, X. Yang, X. Li, et al., Complete global and bifurcation analysis of a stoichiometricpredator-prey model, J. Dyn. Differ. Equ., 30 (2018), 447-472. |
| [23] | G. A. Van Voorn, B. W. Kooi and M. P. Boer, Ecological consequences of global bifurcations in somefood chain models, Math. Biosci., 226 (2010), 120-133. |
| [24] | A. Peace, H. Wang and Y. Kuang, Dynamics of a producer-grazer model incorporating the effects of excess food nutrient content on grazers growth, Bull. Math. Biol., 76 (2014), 2175-2197. |
| [25] |
X. Yang, X. Li, H. Wang, et al., Stability and bifurcation in a stoichiometric producer-grazermodel with knife edge, SIAM J. Appl. Dyn. Syst., 15 (2016), 2051-2077. doi: 10.1137/15M1023610

|
| [26] | M. G. Neubert and H. Caswell, Alternatives to resilience for measuring the responses of ecologicalsystems to perturbations, Ecology, 78 (1997), 653-665. |
| [27] | S. Rinaldi, S. Muratori and Y. Kuznetsov, Multiple attractors, catastrophes and chaos in seasonallyperturbed predator-prey communities, Bull Math. Biol., 55 (1993), 15-35. |
| [28] | S. M. Henson and J. M. Cushing, The effect of periodic habitat fluctuations on a nonlinear insectpopulation model, J. Math. Biol., 36 (1997), 201-226. |
| [29] | M. N. Hassan, L. Asik, J. Kulik, et al., Environmental seasonality on predator-preysystems under nutrient and toxicant constraints, J. Theor. Biol., 480 (2019), 71-80. |
| [30] | R. Vesipa and L. Ridolfi, Impact of seasonal forcing on reactive ecological systems, J. Theor. Biol., 419 (2017), 23-35. |
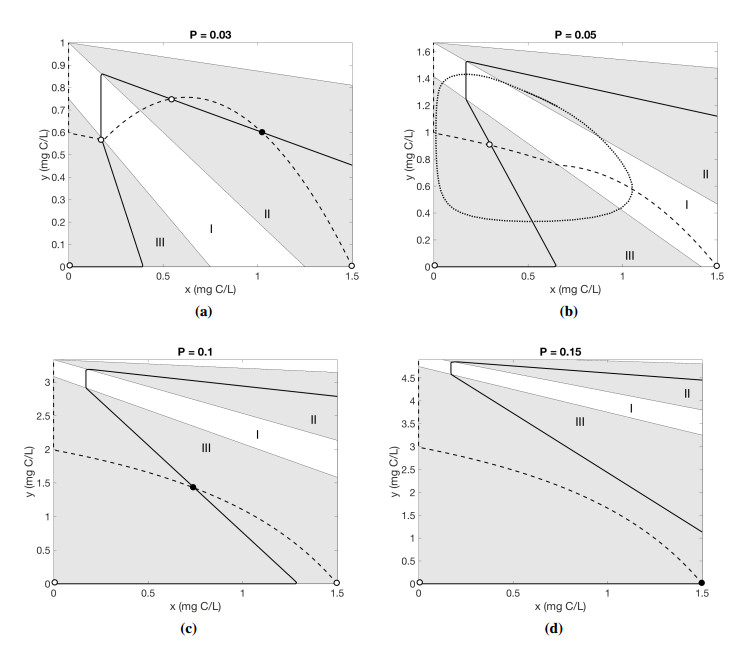
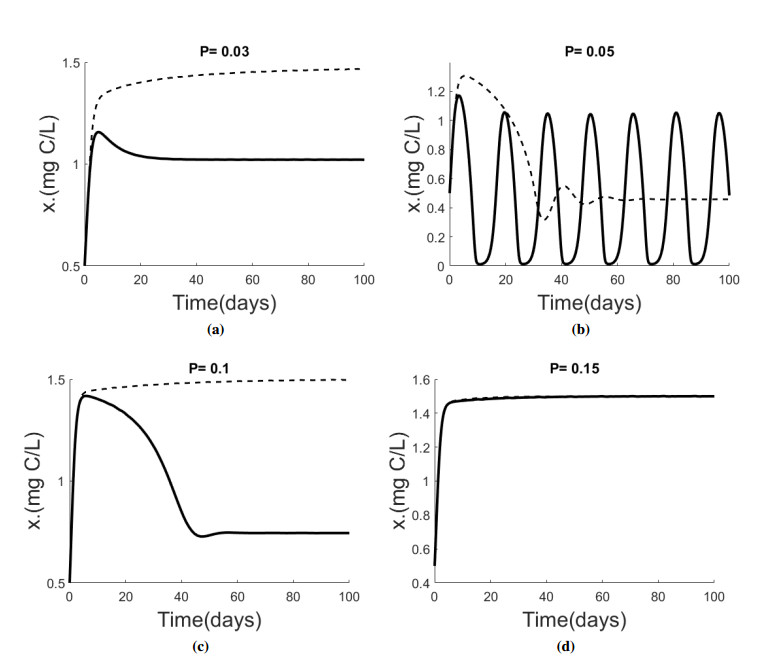
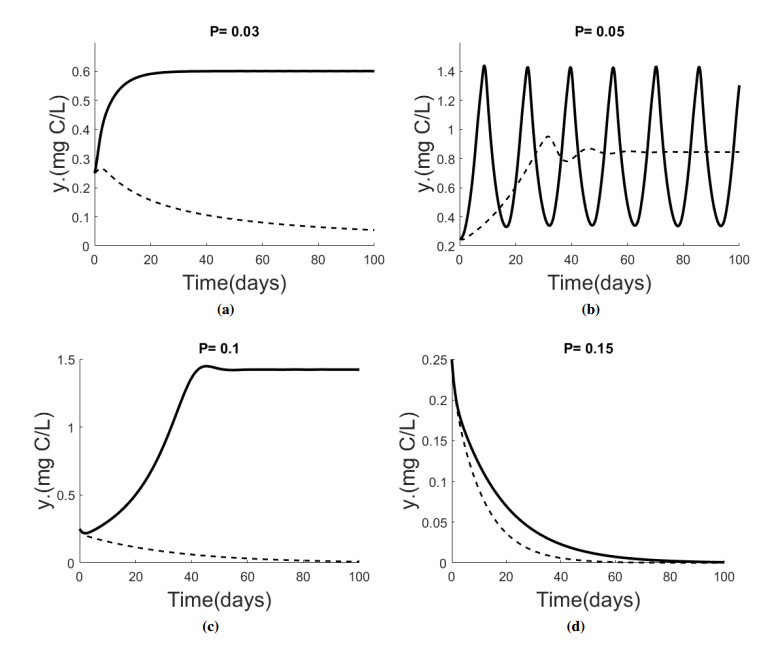
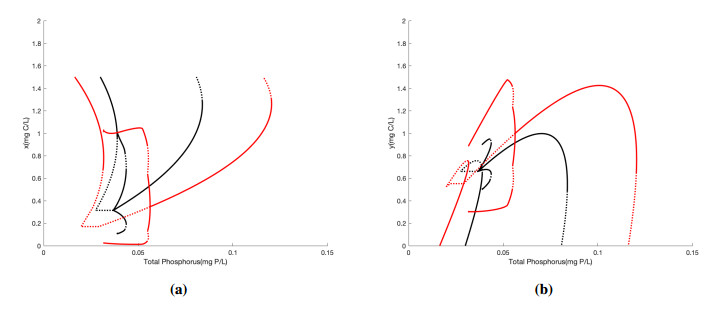
Figures(5) / Tables(1)

Md Nazmul Hassan, Angela Peace. Mechanistically derived Toxicant-mediated predator-prey model under Stoichiometric constraints[J]. Mathematical Biosciences and Engineering, 2020, 17(1): 349-365. doi: 10.3934/mbe.2020019







 DownLoad:
DownLoad: