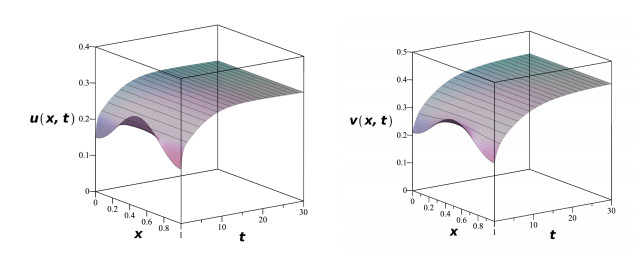
In this paper, we study a system of nonlinear partial differential equations that models the population dynamics of two competitive species both under Allee effects. The consideration of the model includes Logistic growth with Allee effects, Lotka-Volterra competition, diffusion, initial density and boundary conditions on the habitat. In the reaction-diffusion system, we employ the method of upper and lower solutions to address questions on self-elimination or persistence, as well as permanence or competitive exclusion. Specific conditions on biological parameters are explicitly given for extinction, coexistence and competitive exclusion of the species under various boundary conditions. Numerical simulations for the model are demonstrated to illustrate our results from mathematical analysis.
Citation: Yaw Chang, Wei Feng, Michael Freeze, Xin Lu, Charles Smith. Elimination, permanence, and exclusion in a competition model under Allee effects[J]. AIMS Mathematics, 2023, 8(4): 7787-7805. doi: 10.3934/math.2023391
In this paper, we study a system of nonlinear partial differential equations that models the population dynamics of two competitive species both under Allee effects. The consideration of the model includes Logistic growth with Allee effects, Lotka-Volterra competition, diffusion, initial density and boundary conditions on the habitat. In the reaction-diffusion system, we employ the method of upper and lower solutions to address questions on self-elimination or persistence, as well as permanence or competitive exclusion. Specific conditions on biological parameters are explicitly given for extinction, coexistence and competitive exclusion of the species under various boundary conditions. Numerical simulations for the model are demonstrated to illustrate our results from mathematical analysis.

| [1] |
W. C. Allee, E. S. Bowen, Studies in animal aggregations: mass protection against aolloidal silver among goldfishes, J. Exp. Biol., 61 (1932), 185–207. https://doi.org/10.1002/jez.1400610202 doi: 10.1002/jez.1400610202

|
| [2] |
J. Blat, K. J. Brown, Bifurcation of steady-state solutions in predator-prey and competition systems, Proceedings of the Royal Society of Edinburgh, 97 (1984), 21–34. https://doi.org/10.1017/S0308210500031802 doi: 10.1017/S0308210500031802
|
| [3] |
R. S. Cantrell, C. Cosner, V. Hutson, Permanence in ecological systems with spatial heterogeneity, Proceedings of the Royal Society of Edinburgh, 123 (1993), 533–559. https://doi.org/10.1017/S0308210500025877 doi: 10.1017/S0308210500025877
|
| [4] |
Y. Chang, W. Feng, M. Freeze, X. Lu, Permanence and coexistence in a diffusive complex ratio-dependent food chain, Int. J. Dyn. Control, 3 (2015), 262–274. https://doi.org/10.1007/s40435-014-0131-4 doi: 10.1007/s40435-014-0131-4
|
| [5] | L. Edelstein-Keshet, Mathematical models in biology, Philadelphia: Society for Industrial and Applied Mathematics, 2005. https://doi.org/10.1137/1.9780898719147 |
| [6] |
W. Feng, Coexistence, stability, and limiting behavior in a one-predator-two-prey model, J. Math. Anal. Appl., 179 (1993), 592–609. https://doi.org/10.1006/jmaa.1993.1371 doi: 10.1006/jmaa.1993.1371
|
| [7] |
W. Feng, Permanence effect in a three-species food chain model, Appl. Anal., 54 (1994), 195–209. https://doi.org/10.1080/00036819408840277 doi: 10.1080/00036819408840277
|
| [8] |
W. Feng, Competitive exclusion and persistence in models of resource and sexual competition, J. Math. Biol., 35 (1997), 683–694. https://doi.org/10.1007/s002850050071 doi: 10.1007/s002850050071
|
| [9] |
W. Feng, M. Freeze, X. Lu, On competition models under Allee effect: asymptotic behavior and traveling waves, Commun. Pure Appl. Anal., 19 (2020), 5609–5626. http://dx.doi.org/10.3934/cpaa.2020256 doi: 10.3934/cpaa.2020256
|
| [10] |
W. Feng, X. Lu, Some coexistence and extinction results for a 3-species ecological system, Differ. Integral Equ., 8 (1995), 617–626. http://dx.doi.org/10.57262/die/1369316510 doi: 10.57262/die/1369316510
|
| [11] |
W. Feng, X. Lu, Harmless delays for permanence in a class of population models with diffusion effects, J. Math. Anal. Appl., 206 (1997), 547–566. https://doi.org/10.1006/jmaa.1997.5265 doi: 10.1006/jmaa.1997.5265
|
| [12] |
W. Feng, W. Ruan, Coexistence, permanence, and stability in a three species competition model, Acta Math. Appl. Sinica (English Ser.), 12 (1996), 443–446. https://doi.org/10.1007/BF02029074 doi: 10.1007/BF02029074
|
| [13] | A. Leung, Nonlinear systems of partial differential equations: applications to life and physical sciences, Singapore: World Scientific, 2009. https://doi.org/10.1142/7353 |
| [14] |
H. Liu, Y. Ye, Y. Wei, W. Ma, M. Ma, K. Zhang, Pattern formation in a reaction-diffusion predator-prey model with weak Allee effect and delay, Complexity, 2019 (2019), 6282958. https://doi.org/10.1155/2019/6282958 doi: 10.1155/2019/6282958
|
| [15] |
X. Lu, W. Feng, Dynamics and numerical simulations of food-chain populations, Appl. Math. Comput., 65 (1994), 335–344. https://doi.org/10.1016/0096-3003(94)90186-4 doi: 10.1016/0096-3003(94)90186-4
|
| [16] | C. V. Pao, Nonlinear parabolic and elliptic equations, New York: Plenum Press, 1992. https://doi.org/10.1007/978-1-4615-3034-3 |
| [17] |
J. Shi, R. Shivaji, Persistence in reaction diffusion models with weak Allee effect, J. Math. Biol., 52 (2006), 807–829. http://dx.doi.org/10.1007/s00285-006-0373-7 doi: 10.1007/s00285-006-0373-7
|
| [18] |
G. Wang, X. Liang, F. Wang, The competitive dynamics of populations subject to an Allee effect, Ecol. Modell., 124 (1999), 183–192. https://doi.org/10.1016/S0304-3800(99)00160-X doi: 10.1016/S0304-3800(99)00160-X
|
| [19] |
L. Zhou, C. V. Pao, Asymptotic behavior of a competition-diffusion system in population dynamics, Nonlinear Anal.: Theory Method Appl., 6 (1982), 1163–1184. https://doi.org/10.1016/0362-546X(82)90028-1 doi: 10.1016/0362-546X(82)90028-1
|
| [20] |
S. Zhou, C. Liu, G. Wang, The competitive dynamics of metapopulationss subject to the Allee-like effect, Theor. Popul. Biol., 65 (2004), 29–37. https://doi.org/10.1016/j.tpb.2003.08.002 doi: 10.1016/j.tpb.2003.08.002
|
| [21] |
S. Zhou, Y. Liu, G. Wang, The stability of predator-prey systems subject to the Allee effects, Theor. Popul. Biol., 67 (2005), 23–31. https://doi.org/10.1016/j.tpb.2004.06.007 doi: 10.1016/j.tpb.2004.06.007
|




Figures(5)

Yaw Chang, Wei Feng, Michael Freeze, Xin Lu, Charles Smith. Elimination, permanence, and exclusion in a competition model under Allee effects[J]. AIMS Mathematics, 2023, 8(4): 7787-7805. doi: 10.3934/math.2023391







 DownLoad:
DownLoad: