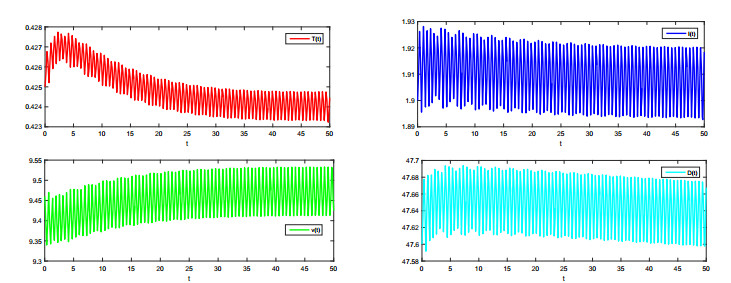
In this paper, a dynamic model of Middle East Respiratory Syndrome Coronavirus (MERS-CoV) with periodic coefficients is proposed and studied. By using the continuation theorem of the coincidence degree theory, we obtain some sufficient conditions for the existence of positive periodic solutions of the model. The periodic model degenerates to an autonomous case, and our conditions can be degenerated to the basic reproductive number $ R_0 > 1 $. Finally, we give some numerical simulations to illustrate our main theoretical results.
Citation: Tuersunjiang Keyoumu, Wanbiao Ma, Ke Guo. Existence of positive periodic solutions for a class of in-host MERS-CoV infection model with periodic coefficients[J]. AIMS Mathematics, 2022, 7(2): 3083-3096. doi: 10.3934/math.2022171
In this paper, a dynamic model of Middle East Respiratory Syndrome Coronavirus (MERS-CoV) with periodic coefficients is proposed and studied. By using the continuation theorem of the coincidence degree theory, we obtain some sufficient conditions for the existence of positive periodic solutions of the model. The periodic model degenerates to an autonomous case, and our conditions can be degenerated to the basic reproductive number $ R_0 > 1 $. Finally, we give some numerical simulations to illustrate our main theoretical results.

| [1] |
A. M. Zaki, S. van Boheemen, T. M. Bestebroer, A. D. M. E. Osterhauset, R. A. M. Fouchier, Isolation of a novel coronavirus from a man with pneumonia in saudi arabia, New Engl. J. Med., 367 (2012), 1814–1820. doi: 10.1056/NEJMoa1211721. doi: 10.1056/NEJMoa1211721

|
| [2] |
A. Assiri, J. A. Al-Tawfiq, A. A. Al-Rabeeah, F. A. Al-Rabiah, S. Al-Hajjar, A. Al-Barrak, et al., Epidemiological, demographic, and clinical characteristics of 47 cases of Middle East respiratory syndrome coronavirus disease from Saudi Arabia: A descriptive study, Lancet Infect. Dis., 13 (2013), 752–761. doi: 10.1016/S1473-3099(13)70204-4. doi: 10.1016/S1473-3099(13)70204-4
|
| [3] |
M. S. Nassar, M. A. Bakhrebah, S. A. Meo, M. S. Alsuabeyl, W. A. Zaheret, Middle east respiratory syndrome coronavirus (MERS-CoV) infection: Epidemiology, pathogenesis and clinical characteristics, Eur. Rev. Med. Pharmaco., 22 (2018), 4956–4961. doi: 10.26355/eurrev_201808_15635. doi: 10.26355/eurrev_201808_15635
|
| [4] |
A. M. Alnuqaydan, A. G. Almutary, A. Sukamaran, B. T. W. Yang, X. T. Lee, W. X. Lim, et al., Middle east respiratory syndrome (MERS) virus-pathophysiological axis and the current treatment strategies, AAPS PharmSciTech, 22 (2021), 173. doi: 10.1208/s12249-021-02062-2. doi: 10.1208/s12249-021-02062-2
|
| [5] | World Health Organization, Middle east respiratory syndrome coronavirus (MERS-CoV), 2019. |
| [6] |
Y. M. Arabi, H. H. Balkhy, F. G. Hayden, A. Bouchama, T. Luke, J. K. Baillie, et al., Middle east respiratory syndrome, New Engl. J. Med., 376 (2017), 584–594. doi: 10.1056/NEJMsr1408795. doi: 10.1056/NEJMsr1408795
|
| [7] |
V. S. Raj, H. Mou, S. L. Smits, D. H. W. Dekkers, M. A. Müller, R. Dijkman, et al., Dipeptidyl peptidase 4 is a functional receptor for the emerging human coronavirus-EMC, Nature, 495 (2013), 251–254. doi: 10.1038/nature12005. doi: 10.1038/nature12005
|
| [8] |
G. Lu, Y. Hu, Q. Wang, J. Qi, F. Gao, Y. Li, et al., Molecular basis of binding between novel human coronavirus MERS-CoV and its receptor CD26, Nature, 500 (2013), 227–231. doi: 10.1038/nature12328. doi: 10.1038/nature12328
|
| [9] |
M. A. Nowak, C. R. M. Bangham, Population dynamics of immune responses to persistent virus, Science, 272 (1997), 74–79. doi: 10.1126/science.272.5258.74. doi: 10.1126/science.272.5258.74
|
| [10] |
A. U. Neumann, N. P. Lam, H. Dahari, D. R. Gretch, T. E. Wiley, T. J. Layden, et al., Hepatitis C viral dynamics in vivo and the antiviral efficacy of interferon-$\alpha$ therapy, Science, 282 (1998), 103–107. doi: 10.1126/science.282.5386.103. doi: 10.1126/science.282.5386.103
|
| [11] |
A. S. Perelson, P. W. Nelson, Mathematical analysis of HIV-1 dynamics in vivo, SIAM Rev., 41 (1999), 3–44. doi: 10.1137/S0036144598335107. doi: 10.1137/S0036144598335107
|
| [12] |
S. Tang, W. Ma, P. Bai, A novel dynamic model describing the spread of the MERS-CoV and the expression of dipeptidyl peptidase 4, Comput. Math. Method. M., 2017 (2017), 5285810. doi: 10.1155/2017/5285810. doi: 10.1155/2017/5285810
|
| [13] |
A. N. Chatterjee, F. Al Basir, A model for SARS-CoV-2 infection with treatment, Comput. Math. Method. M., 2020 (2020), 1352982. doi: 10.1155/2020/1352982. doi: 10.1155/2020/1352982
|
| [14] |
P. D. Leenheer, Within-host virus models with periodic antiviral therapy, Bull. Math. Biol., 71 (2009), 189–210. doi: 10.1007/s11538-008-9359-5. doi: 10.1007/s11538-008-9359-5
|
| [15] |
Y. Yang, Y. Xiao, Threshold dynamics for an HIV model in periodic environments, J. Math. Anal. Appl., 361 (2010), 59–68. doi: 10.1016/j.jmaa.2009.09.012. doi: 10.1016/j.jmaa.2009.09.012
|
| [16] | R. E. Gaines, J. L. Mawhin, Coincidence degree and nonlinear differential equations, Berlin: Springer-Verlag, 1977. doi: 10.1007/BFb0089537. |
| [17] | Y. Li, Periodic solutions of a periodic delay predator-prey system, P. Am. Math. Soc., 127 (1999), 1331–1335. doi: S0002-9939(99)05210-7. |
| [18] | Y. Li, Y. Kuang, Periodic solutions of periodic delay Lotka-Volterra equations and systems, J. Math. Anal. Appl., 255 (2001), 260–280. doi: 10.1006jmaa.2000.7248. |
| [19] |
M. Fan, K. Wang, Periodicity in a delayed ratio-dependent predator-prey system, J. Math. Anal. Appl., 262 (2001), 179–190. doi: 10.1006/jmaa.2001.7555. doi: 10.1006/jmaa.2001.7555
|
| [20] |
D. Bai, J. Yu, M. Fan, Y. Kang, Dynamics for a non-autonomous predator-prey system with generalist predator, J. Math. Anal. Appl., 485 (2020), 123820. doi: 10.1016/j.jmaa.2019.123820. doi: 10.1016/j.jmaa.2019.123820
|
| [21] |
Z. Bai, Y. Zhou, T. Zhang, Existence of multiple periodic solutions for an SIR model with seasonality, Nonlinear Anal., 74 (2011), 3548–3555. doi: 10.1016/j.na.2011.03.008. doi: 10.1016/j.na.2011.03.008
|
| [22] |
T. Zhang, J. Liu, Z. Teng, Existence of positive periodic solutions of an SEIR model with periodic coefficients, Appl. Math. CZECH, 57 (2012), 601–616. doi: 10.1007/s10492-012-0036-5. doi: 10.1007/s10492-012-0036-5
|
| [23] |
H. Zhao, L. Sun, Periodic oscillatory and global attractivity for chemostat model involving distributed delays, Nonlinear Anal.-Real, 7 (2006), 385–394. doi: 10.1016/j.nonrwa.2005.03.007. doi: 10.1016/j.nonrwa.2005.03.007
|
| [24] |
P. Amster, G. Robledo, D. Sepúlveda, Existence of $\omega$-periodic solutions for a delayed chemostat with periodic inputs, Nonlinear Anal.-Real, 55 (2020), 103134. doi: 10.1016/j.nonrwa.2020.103134. doi: 10.1016/j.nonrwa.2020.103134
|
Figures(1)

Tuersunjiang Keyoumu, Wanbiao Ma, Ke Guo. Existence of positive periodic solutions for a class of in-host MERS-CoV infection model with periodic coefficients[J]. AIMS Mathematics, 2022, 7(2): 3083-3096. doi: 10.3934/math.2022171







 DownLoad:
DownLoad: