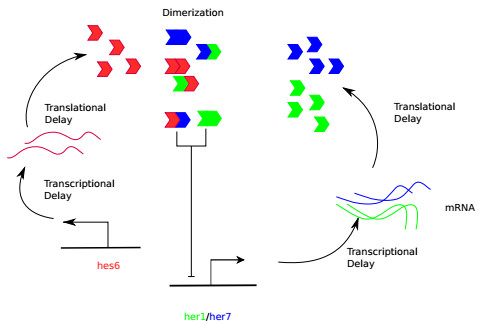
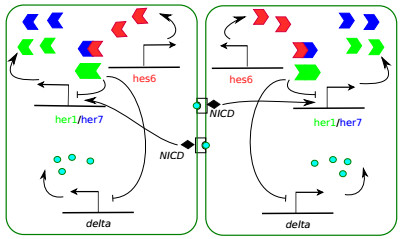
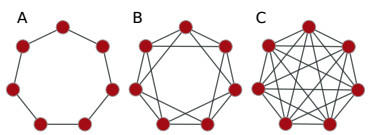
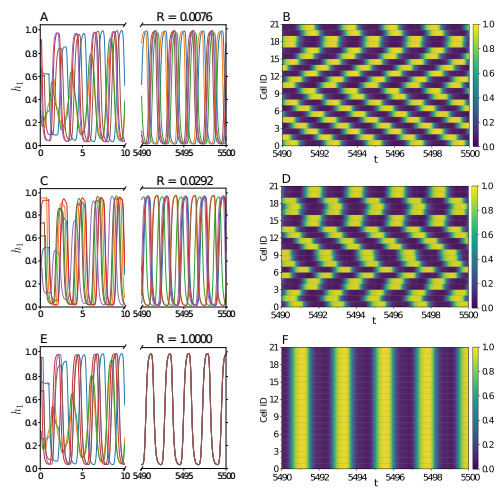
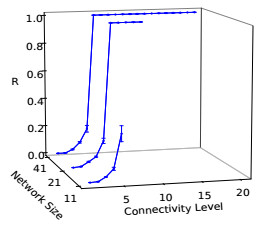
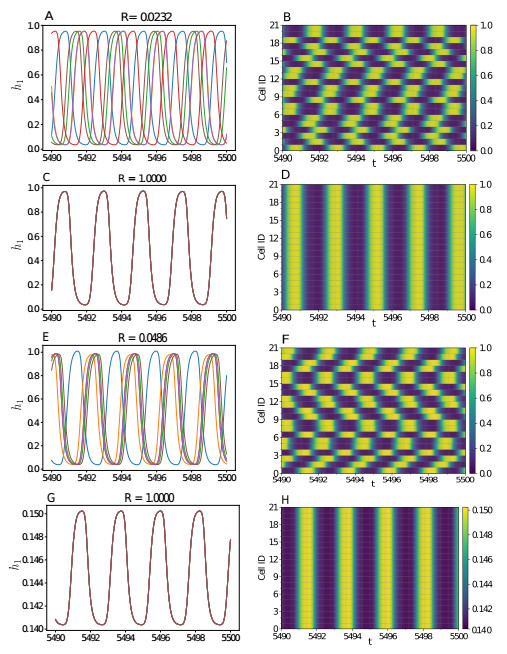
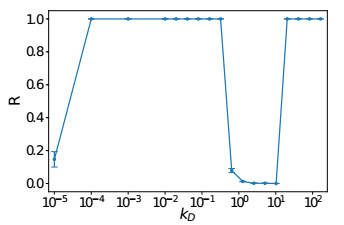
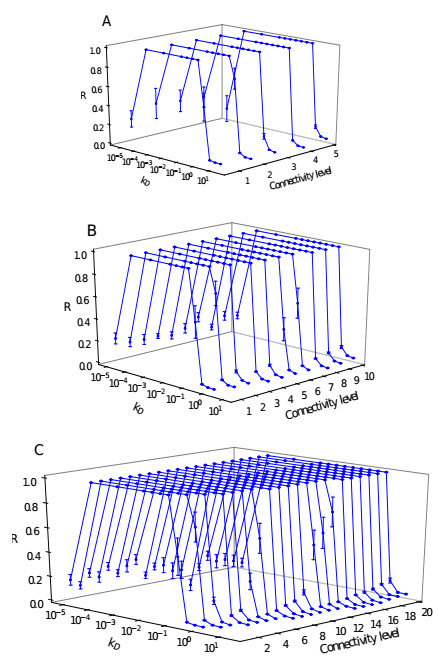
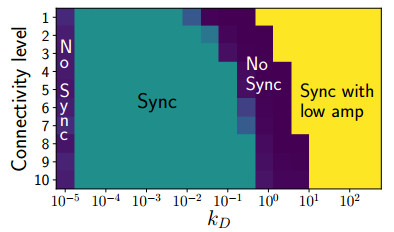
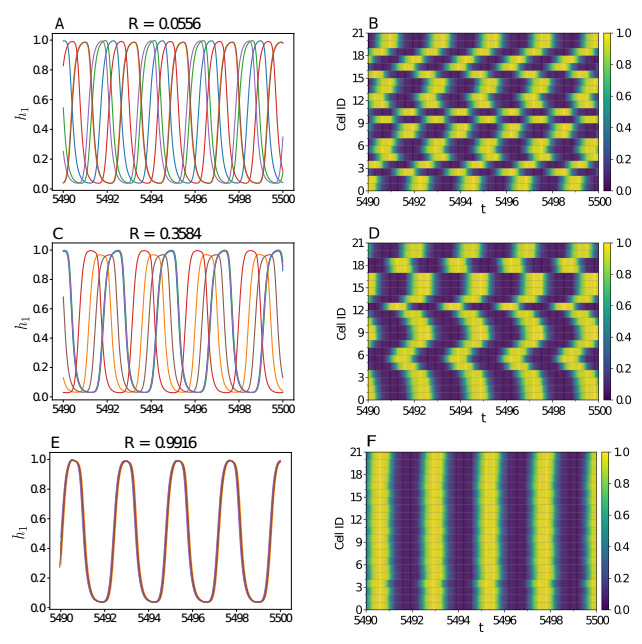
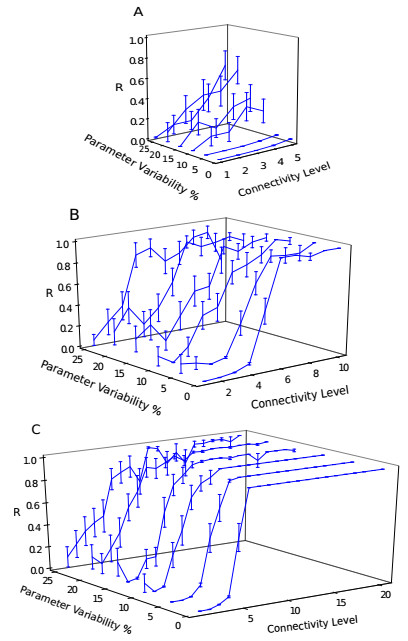
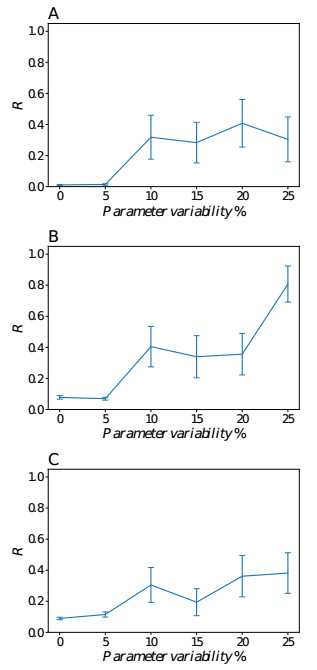
Somitogenesis is the process by means of which a tissue known as presomitic mesoderm (PSM) is segmented in blocks of cells, called somites, along the anterior-posterior axis of the developing embryo in segmented animals. In vertebrates, somites give rise to axial skeleton, cartilage, tendons, skeletal muscle, and dermis. Somite formation occurs periodically, and this periodicity is driven by a genetic oscillator that operates within PSM cells and is known as the segmentation clock. The correct synchronization of the segmentation clock among PSM cells is essential for somitogenesis to develop normally. When synchronization is disrupted, somites form irregularly and, in consequence, the tissues that originate from them show clear malformations. In this work, based in a model for zebrafish segmentation clock, we investigate by means of a mathematical modeling approach, how PSM-cell synchronization is affected by factors like: the size of PSM-cell networks, the amount of cell-to-cell interactions per PSM cell, the strength of these interactions, and the inherent variability among PSM cells. Interestingly we found that very small PSM-cell networks are unable to synchronize. Moreover, the effect of decreasing the strength of interactions among PSM cells is corrected by increasing the network connectivity-level, and a moderated level of variability among cells can have a positive effect on synchronization, specially in large networks.
Citation: Jesús Pantoja-Hernández, Moisés Santillán. Segmentation-clock synchronization in circular-lattice networks of embryonic presomitic-mesoderm cells[J]. AIMS Mathematics, 2021, 6(6): 5817-5836. doi: 10.3934/math.2021344
Somitogenesis is the process by means of which a tissue known as presomitic mesoderm (PSM) is segmented in blocks of cells, called somites, along the anterior-posterior axis of the developing embryo in segmented animals. In vertebrates, somites give rise to axial skeleton, cartilage, tendons, skeletal muscle, and dermis. Somite formation occurs periodically, and this periodicity is driven by a genetic oscillator that operates within PSM cells and is known as the segmentation clock. The correct synchronization of the segmentation clock among PSM cells is essential for somitogenesis to develop normally. When synchronization is disrupted, somites form irregularly and, in consequence, the tissues that originate from them show clear malformations. In this work, based in a model for zebrafish segmentation clock, we investigate by means of a mathematical modeling approach, how PSM-cell synchronization is affected by factors like: the size of PSM-cell networks, the amount of cell-to-cell interactions per PSM cell, the strength of these interactions, and the inherent variability among PSM cells. Interestingly we found that very small PSM-cell networks are unable to synchronize. Moreover, the effect of decreasing the strength of interactions among PSM cells is corrected by increasing the network connectivity-level, and a moderated level of variability among cells can have a positive effect on synchronization, specially in large networks.

| [1] |
O. V. Popovych, C. Hauptmann, P. A. Tass, Control of neuronal synchrony by nonlinear delayed feedback, Biol. Cybern., 95 (2006), 69–85. doi: 10.1007/s00422-006-0066-8

|
| [2] | S. Strogatz, Sync, Hyperion, New York, 1 edition, 2003. |
| [3] | P. Arkady, Synchronization, Boris Chirikov, Predrag Cvitanovic, Frank Moss, Harry Swinney, New York, 1 edition, 2001. |
| [4] |
L. A. Rohde, C. P. Heisenberg, Zebrafish gastrulation: Cell movements, signals, and mechanisms, Int. Rev. Cytol., 261 (2007), 159–192. doi: 10.1016/S0074-7696(07)61004-3
|
| [5] | S. Gilbert, Developmental biology, Sinauer Associates, Inc., Publishers, Sunderland, Massachusetts, 2016. |
| [6] | T. W. Sadler, Langman's medical embryology, Wolters Kluwer Health/Lippincott Williams & Wilkins, Philadelphia, 2012. |
| [7] | B. Carlson, Human embryology and developmental biology, Elsevier/Saunders, Philadelphia, Pa, 2014. |
| [8] | Y. Harima, R. Kageyama, Oscillatory links of fgf signaling and hes7 in the segmentation clock. Curr. Opin. Genet. Dev., 23 (2013), 484–490. |
| [9] | P. F. Giampietro, S. L. Dunwoodie, K. Kusumi, O. Pourquié, O. Tassy, A. C. Offiah, et al. Progress in the understanding of the genetic etiology of vertebral segmentation disorders in humans, Ann. NY Acad. Sci., 1151 (2008), 38–67. |
| [10] |
M. L. Dequéant, O. Pourquié, Segmental patterning of the vertebrate embryonic axis, Nat. Rev. Genet., 9 (2008), 370–382. doi: 10.1038/nrg2320
|
| [11] | M. Maroto, R. A. Bone, J. K. Dale, Somitogenesis, Development, 139 (2012), 2453–2456. |
| [12] |
J. Cooke, E. C. Zeeman, A clock and wavefront model for control of the number of repeated structures during animal morphogenesis, J. Theor. Biol., 58 (1976), 455–476. doi: 10.1016/S0022-5193(76)80131-2
|
| [13] |
I. Palmeirim, D. Henrique, D. Ish-Horowicz, O. Pourquié, Avian hairy gene expression identifies a molecular clock linked to vertebrate segmentation and somitogenesis, Cell, 91 (1997), 639–648. doi: 10.1016/S0092-8674(00)80451-1
|
| [14] |
M.-L. Dequeant, E. Glynn, K. Gaudenz, M. Wahl, J. Chen, A. Mushegian, et al. A complex oscillating network of signaling genes underlies the mouse segmentation clock, Science, 314 (2006), 1595–1598. doi: 10.1126/science.1133141
|
| [15] |
C. Gomez, E. M. Özbudak, J. Wunderlich, D. Baumann, J. Lewis, O. Pourquié, Control of segment number in vertebrate embryos, Nature, 454 (2008), 335–339. doi: 10.1038/nature07020
|
| [16] | H. Y. Wang, Y. X. Huang, L. H. Zheng, Y. L. Bao, L. G. Sun, Y. Wu, et al. Modelling coupled oscillations in the notch, wnt, and FGF signaling pathways during somitogenesis: A comprehensive mathematical model, Comput. Intel. Neurosc., 2015 (2015), 1–16. |
| [17] |
J. Lewis, Autoinhibition with transcriptional delay, Curr. Biol., 13 (2003), 1398–1408. doi: 10.1016/S0960-9822(03)00534-7
|
| [18] | A. B. Webb, I. M. Lengyel, D. J. Jörg, G. Valentin, F. Jülicher, L. G. Morelli, et al. Persistence, period and precision of autonomous cellular oscillators from the zebrafish segmentation clock, eLife, 5 (2016), e08438. |
| [19] | E. M. Özbudak, J. Lewis, Notch signalling synchronizes the zebrafish segmentation clock but is not needed to create somite boundaries, PLoS Genet., 4 (2008), e15. |
| [20] |
Y.-J. Jiang, B. L. Aerne, L. Smithers, C. Haddon, D. Ish-Horowicz, J. Lewis, Notch signalling and the synchronization of the somite segmentation clock, Nature, 408 (2000), 475–479. doi: 10.1038/35044091
|
| [21] |
J. Lewis, A. Hanisch, M. Holder, Notch signaling, the segmentation clock, and the patterning of vertebrate somites, J. Biol., 8 (2009), 1–7. doi: 10.1186/jbiol111
|
| [22] |
W. R. Gordon, K. L. Arnett, S. C. Blacklow, The molecular logic of notch signaling - a structural and biochemical perspective, J. Cell Sci., 121 (2008), 3109–3119. doi: 10.1242/jcs.035683
|
| [23] | R. Kopan, Current topics in development biology: Notch Signaling, Academic, San Diego, CA, 2010. |
| [24] | E. R. Andersson, U. Lendahl, Therapeutic modulation of notch signalling — are we there yet? Nat. Rev. Drug Discov., 13 (2014), 357–378. |
| [25] | S. J. Bray, Notch signalling: a simple pathway becomes complex, Nat. Rev. Mol. Cell Bio., 7 (2006), 678–689. |
| [26] |
H. B. Tiedemann, E. Schneltzer, S. Zeiser, I. Rubio-Aliaga, W. Wurst, J. Beckers, et al. Cell-based simulation of dynamic expression patterns in the presomitic mesoderm, J. Theor. Biol., 248 (2007), 120–129. doi: 10.1016/j.jtbi.2007.05.014
|
| [27] |
I. H. Riedel-Kruse, C. Muller, A. C. Oates, Synchrony dynamics during initiation, failure, and rescue of the segmentation clock, Science, 317 (2007), 1911–1915. doi: 10.1126/science.1142538
|
| [28] |
K. Horikawa, K. Ishimatsu, E. Yoshimoto, S. Kondo, H. Takeda, Noise-resistant and synchronized oscillation of the segmentation clock, Nature, 441 (2006), 719–723. doi: 10.1038/nature04861
|
| [29] |
K. Uriu, Y. Morishita, Y. Iwasa, Random cell movement promotes synchronization of the segmentation clock, P.Nat. Acad. Sci., 107 (2010), 4979–4984. doi: 10.1073/pnas.0907122107
|
| [30] | O. Cinquin, Repressor dimerization in the zebrafish somitogenesis clock, PLoS Comput. Biol., 3 (2007), e32. |
| [31] | C. A. Henry, M. K. Urban, K. K. Dill, J. P. Merlie, M. F. Page, C. B. Kimmel, et al. Two linked hairy/Enhancer of split-related zebrafish genes, her1 and her7, function together to refine alternating somite boundaries, Development, 129 (2002), 3693–3704. |
| [32] | A. C. Oates, R. K. Ho, Hairy/E(spl)-related (Her) genes are central components of the segmentation oscillator and display redundancy with the Delta/Notch signaling pathway in the formation of anterior segmental boundaries in the zebrafish, Development, 129 (2002), 2929–2946. |
| [33] | C. Schröter, S. Ares, L. G. Morelli, A. Isakova, K. Hens, D. Soroldoni, et al. Topology and dynamics of the zebrafish segmentation clock core circuit, PLoS Biol., 10 (2012), e1001364. |
| [34] |
A. Ay, S. Knierer, A. Sperlea, J. Holland, E. M. Ozbudak, Short-lived her proteins drive robust synchronized oscillations in the zebrafish segmentation clock, Development, 140 (2013), 3244–3253. doi: 10.1242/dev.093278
|
| [35] |
J.S. Griffith, Mathematics of cellular control processes i. negative feedback to one gene, J. Theor. Biol., 20 (1968), 202–208. doi: 10.1016/0022-5193(68)90189-6
|
| [36] |
M. Santillán, On the use of the hill functions in mathematical models of gene regulatory networks, Math. Model. Nat. Pheno., 3 (2008), 85–97. doi: 10.1051/mmnp:2008056
|
| [37] |
D. Gonze, S. Bernard, C. Waltermann, A. Kramer, H. Herzel, Spontaneous synchronization of coupled circadian oscillators, Biophys. J., 89 (2005), 120–129. doi: 10.1529/biophysj.104.058388
|
| [38] |
J. Garcia-Ojalvo, M. B. Elowitz, S. H. Strogatz, Modeling a synthetic multicellular clock: Repressilators coupled by quorum sensing, P. Nat. Acad. Sci., 101 (2004), 10955–10960. doi: 10.1073/pnas.0307095101
|
| [39] | W. Wu, W. J. Zhou, T. P. Chen, Cluster synchronization of linearly coupled complex networks under pinning control, IEEE T. Circuits I, 56 (2009), 829–839. |
| [40] |
T. Dahms, J. Lehnert, E. Schöll, Cluster and group synchronization in delay-coupled networks, Phys. Rev. E, 86 (2012), 016202. doi: 10.1103/PhysRevE.86.016202
|
| [41] |
F. Sorrentino, L. Pecora, Approximate cluster synchronization in networks with symmetries and parameter mismatches, Chaos: An Interdisciplinary Journal of Nonlinear Science, 26 (2016), 094823. doi: 10.1063/1.4961967
|
| [42] |
J. W. Feng, P. Yang, Y. Zhao, Cluster synchronization for nonlinearly time-varying delayed coupling complex networks with stochastic perturbation via periodically intermittent pinning control, Appl. Math. Comput., 291 (2016), 52–68. doi: 10.1016/j.amc.2016.06.030
|
| [43] |
C. Ma, Q. R. Yang, X. Q. Wu, J. A. Lu, Cluster synchronization: From single-layer to multi-layer networks, Chaos: An Interdisciplinary Journal of Nonlinear Science, 29 (2019), 123120. doi: 10.1063/1.5122699
|
| [44] |
G. B. Ermentrout, Oscillator death in populations of "all to all" coupled nonlinear oscillators, Physica D, 41 (1990), 219–231. doi: 10.1016/0167-2789(90)90124-8
|
| [45] |
F. A. Rodrigues, T. K. DM. Peron, P. Ji, J. Kurths, The kuramoto model in complex networks, Physics Reports, 610 (2016), 1–98. doi: 10.1016/j.physrep.2015.10.008
|
| [46] |
J. Gómez-Gardeñes, Y. Moreno, A. Arenas, Synchronizability determined by coupling strengths and topology on complex networks, Phys. Rev. E, 75 (2007), 066106. doi: 10.1103/PhysRevE.75.066106
|
| [47] |
E. A. Delaune, P. François, N. P. Shih, S. L. Amacher, Single-cell-resolution imaging of the impact of notch signaling and mitosis on segmentation clock dynamics, Dev. Cell, 23 (2012), 995–1005. doi: 10.1016/j.devcel.2012.09.009
|
| [48] |
J.-N. Teramae, H. Nakao, G. B. Ermentrout, Stochastic phase reduction for a general class of noisy limit cycle oscillators, Phys. Rev. Lett., 102 (2009), 194102. doi: 10.1103/PhysRevLett.102.194102
|
| [49] |
K. Kotani, I. Yamaguchi, Y. Ogawa, Y. Jimbo, H. Nakao, G. B. Ermentrout, Adjoint method provides phase response functions for delay-induced oscillations, Phys. Rev. Lett., 109 (2012), 044101. doi: 10.1103/PhysRevLett.109.044101
|
Figures(12) / Tables(1)

Jesús Pantoja-Hernández, Moisés Santillán. Segmentation-clock synchronization in circular-lattice networks of embryonic presomitic-mesoderm cells[J]. AIMS Mathematics, 2021, 6(6): 5817-5836. doi: 10.3934/math.2021344







 DownLoad:
DownLoad: