Citation: Kasbawati, Mariani, Nur Erawaty, Naimah Aris. A mathematical study of effects of delays arising from the interaction of anti-drug antibody and therapeutic protein in the immune response system[J]. AIMS Mathematics, 2020, 5(6): 7191-7213. doi: 10.3934/math.2020460

| [1] | J. A. Pedras-Vasconcelos, The Immunogenicity of Therapeutic Proteins-What you Don't Know Can Hurt YOU and the Patient. SBIA REdI Fall, Ed.; U.S. Food and Drug Administration: Silver Spring, MD, USA, 2014. Available from: https://www.fda.gov/files/drugs/published/The-immunogenicity-of-therapeutic-proteins--what-you-don%E2%80%99t-know-can-hurt-YOU-and-the-patient--Fall-2014.pdf. |
| [2] | J. M. Carton, W. R. Strohl, Chapter 4-Protein therapeutics (introduction to biopharmaceuticals), Intro. Biol. Small Mole. Drug Rese. Develop., (2013), 127-159. |
| [3] |
N. Chirmule, V. Jawa, B. Meibohm, Immunogenicity to therapeutic proteins: Impact on PK/PD and Efficacy, AAPS J., 14 (2012), 296-302. doi: 10.1208/s12248-012-9340-y

|
| [4] |
K. D. Ratanji, J. P. Derrick, R. J. Dearman, et al. Immunogenicity of therapeutic proteins: Influence of aggregation, J. Immunotoxicol, 11 (2014), 99-109. doi: 10.3109/1547691X.2013.821564
|
| [5] | A. Kuriakose, N. Chirmule, P. Nair, Immunogenicity of biotherapeutics: Causes and association with posttranslational modifications, J. Immunol. Res., 2016 (2016), 1-18. |
| [6] |
S. Tourdot, T. P. Hickling, Nonclinical immunogenicity risk assessment of therapeutic proteins, Bioanalysis, 11 (2019), 1631-1643. doi: 10.4155/bio-2018-0246
|
| [7] |
G. Shankar, S. Arkin, L. Cocea, et al. Assessment and reporting of the clinical immunogenicity of therapeutic proteins and peptides-harmonized terminology and tactical recommendations, AAPS J., 16 (2014), 658-673. doi: 10.1208/s12248-014-9599-2
|
| [8] |
B. Chen, J. Q. Dong, W. J. Pan, et al. Pharmacokinetics/pharmacodynamics model supported early drug development, Curr Pharm Biotechnol., 13 (2012), 1360-1375. doi: 10.2174/138920112800624436
|
| [9] | A. Smith, H. Manoli, S. Jaw, et al. Unraveling the effect of immunogenicity on the PK/PD, efficacy, and safety of therapeutic proteins, J. Immunol. Res., 2016 (2016), 1-9. |
| [10] | W. H. Boehncke, N. C. Brembilla, Immunogenicity of biologic therapies: Causes and consequences, Exp. Rev. Clin. Immunol., 14 (2018), 513-523. |
| [11] | B. W. Neun, Y. Barenholz, J. Szebeni, et al. Understanding the Role of Anti-PEG Antibodies in the Complement Activation by Doxil in Vitro, Molecule, 23 (2018), 1700. |
| [12] | S. Kathman, T. M. Thway, L. Zhou, et al. Utility of a bayesian mathematical model to predict the impact of immunogenicity on pharmacokinetics of therapeutic proteins, AAPS J., 18 (2016), 424. |
| [13] |
E. M. J. van Brummelen, W. Ros, G. Wolbink, et al. Antidrug antibody formation in oncology: Clinical relevance and challenges, Oncologist, 21 (2016), 1260-1268. doi: 10.1634/theoncologist.2016-0061
|
| [14] | S. Gupta, S. R. Indelicato, V. Jethwa, et al. Recommendations for the design, optimization, and qualification of Cell-Based assays used for the detection of neutralizing antibody responses elicited to biologycal therapeutics, J. Immunol. Methods, 32 (2007), 1-18. |
| [15] | X. Chen, T. P. Hickling, P. Vicini, A mechanistic, multiscale mathematical model of immunogenicity for therapeutic proteins: Part 1-Theoretical model, CPT Pharmacometrics Syst. Pharmacol., 3 (2014), e133. |
| [16] | L. Hamuro, G. S. Tirucherai, S. M. Crawford, et al. Evaluating a multiscale mechanistic model of the immune system to predict human immunogenicity for a biotherapeutic in Phase 1, AAPS J., 21 (2019), 94. |
| [17] |
G. I. Bell, Mathematical model of clonal selection and antibody production, J Theor Biol., 29 (1970), 191-232. doi: 10.1016/0022-5193(70)90019-6
|
| [18] |
H. Y. Lee, D. J. Topham, S. Y. Park, et al. Simulation and prediction of the adaptive immune response to influenza a virus infection, J. Virol., 83 (2009), 7151-7165. doi: 10.1128/JVI.00098-09
|
| [19] |
Z. H. Xu, H. Lee, T. Vu, et al. Populations pharmacokinetics of golimumab in patients with anklosing spondylitis: Impact of body weight and immunogenicity, Int J Clin Pharmacol Ther., 48 (2010), 596-607. doi: 10.5414/CPP48596
|
| [20] | P. L. Bonate, C. Sung, K. Welch, et al. Conditional modeling of antibody titers using A Zero-Inflated poisson random effect model: Application to fabrazyme, J Pharmacokinet Phar., 35 (2009), 449-459. |
| [21] |
X. Chen, T. P. Hickling, E. Kraynov, et al. A mathematical model of the effect of immunogenicity on therapeutic protein pharmacokinetics, AAPS J., 15 (2013), 1141-1154. doi: 10.1208/s12248-013-9517-z
|
| [22] |
J. Foote, C. Milstein, Kinetic maturation of an immune response, Nature, 352 (1991), 530-532. doi: 10.1038/352530a0
|
| [23] | B. S. Alexandra, N. Radhika, H. Paul, Novel Approaches and Strategies for Biologics, Vaccines and Cancer Therapies, Chapter 9-Biobetter Biologics, New York: Academic Press, (2015), 199-217. |
| [24] | Pacific Immunology, Learning Center: Antibody Affinity and Affinity Maturation, 2020., Available from: https://www.pacificimmunology.com/resources/antibody-introduction/antibody-affinity-and-affinity-maturation/. |
| [25] | S. Yi, P. W. Nelson, A. G. Ulsoy, Time-delay sistems: Analysis and control using the Lambert W function, World Scientific, Singapore, 2010. |
| [26] | Kasbawati, A. Y. Gunawan, R. Hertadi, et al. Effects of time delay on the dynamics of a kinetic model of a microbial fermentation process, ANZIAM J., 55 (2014), 336-356. |
| [27] | J. D. Murray, (1990) Mathematical biology, 2Eds., New York: Springer. |
| [28] |
L. F. Shampine, S. Thompson, Solving DDEs in MATLAB, Appl. Numer. Math. 37 (2001), 441-458. doi: 10.1016/S0168-9274(00)00055-6
|
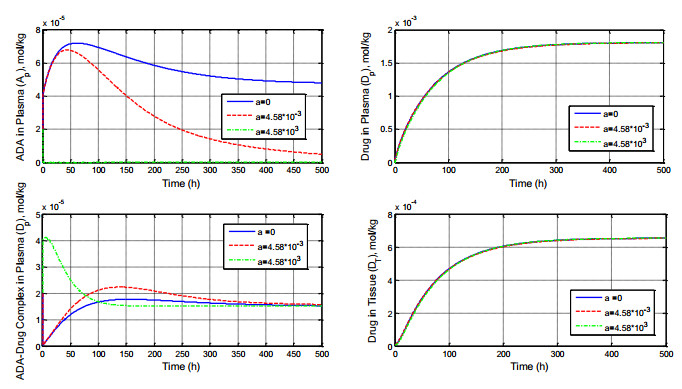
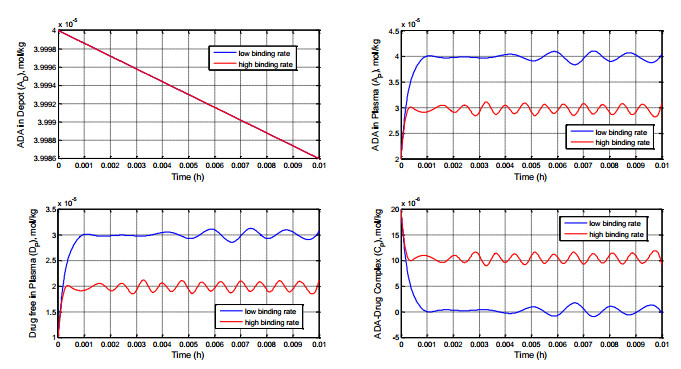
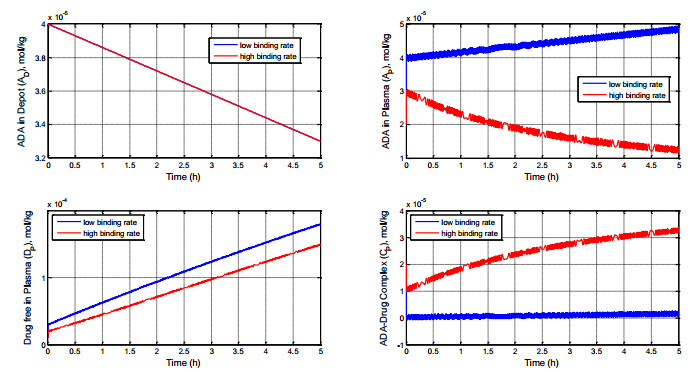
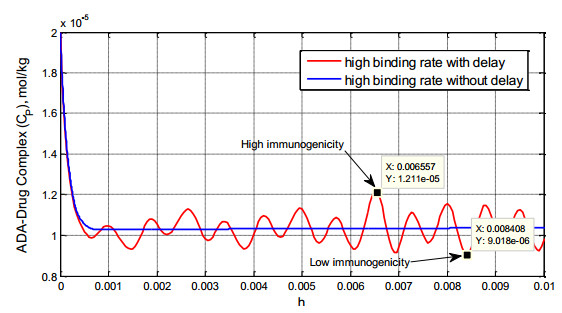
Figures(8) / Tables(1)

Kasbawati, Mariani, Nur Erawaty, Naimah Aris. A mathematical study of effects of delays arising from the interaction of anti-drug antibody and therapeutic protein in the immune response system[J]. AIMS Mathematics, 2020, 5(6): 7191-7213. doi: 10.3934/math.2020460







 DownLoad:
DownLoad: