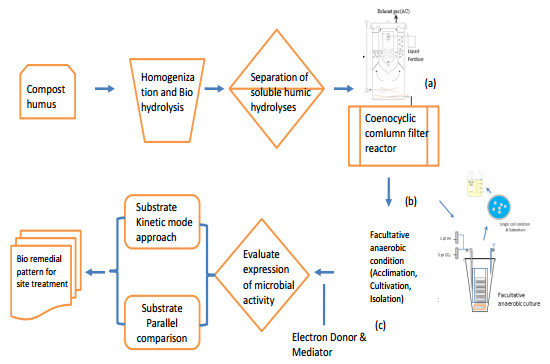
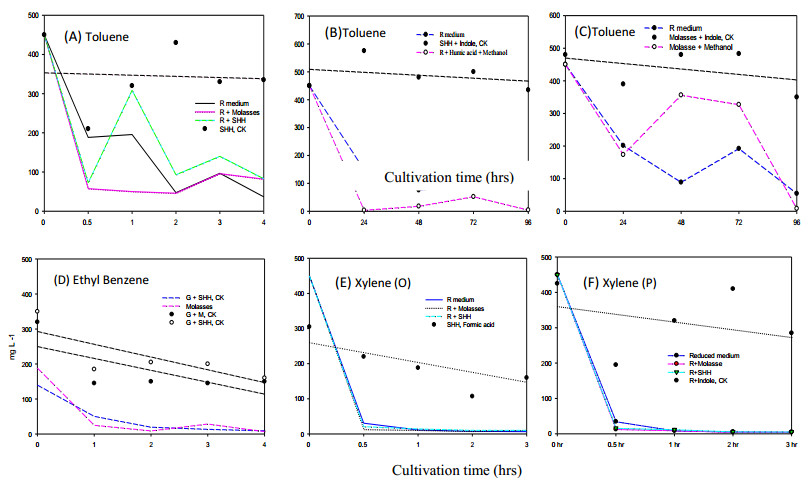
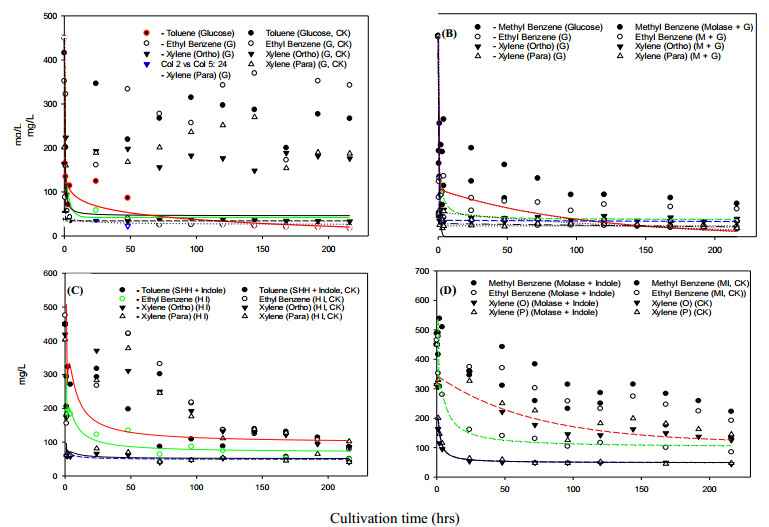
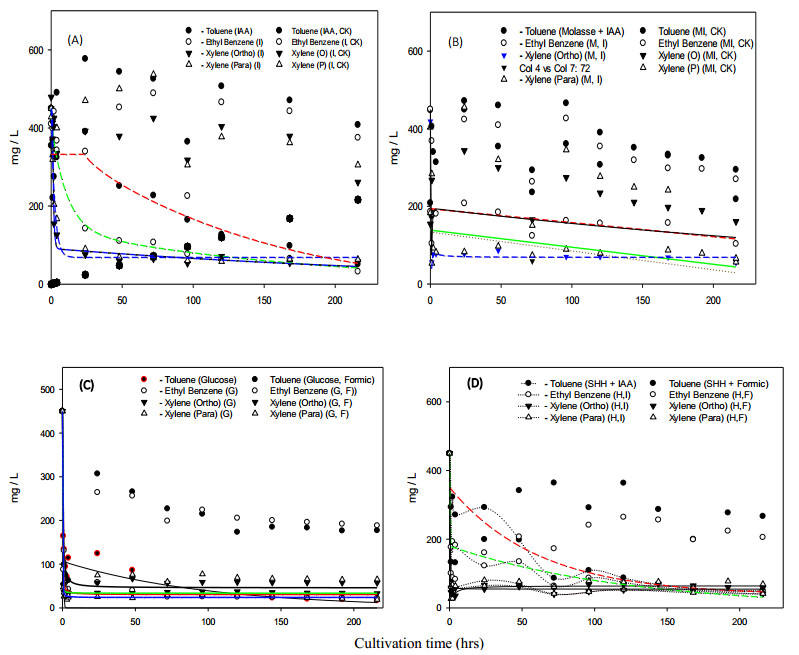
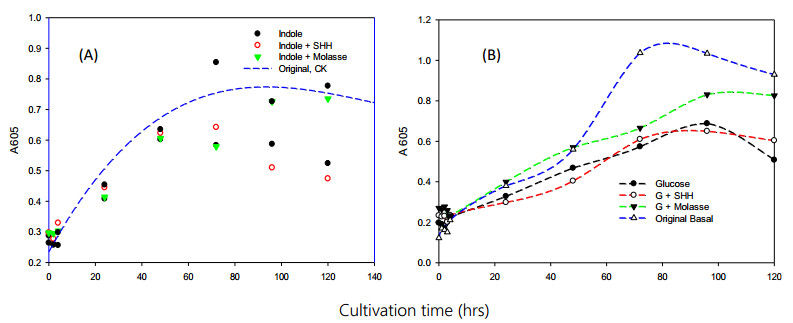
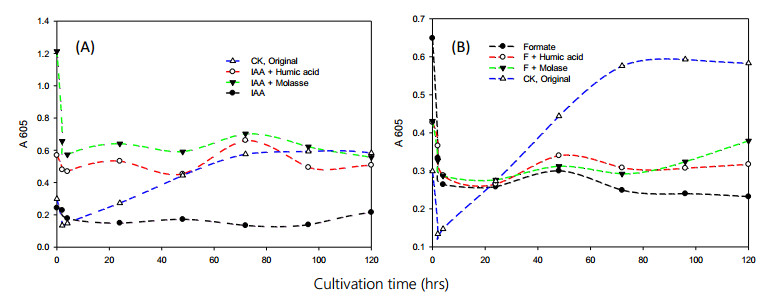
Bioremediation of alkylbenzenes, including toluene, ethylbenzene and xylenes, was performed using fermentable aromatic sources and electron mediators by Bacillus cereus 301 in a limited oxygen state. The fermentation of small fermentable aromatic melanoids from cow manure as soluble humus hydrolysates or sugarcane molasses as saccharine, glucose and limited basal medium was compared. Thus, an evaluation model of exponential decline against a control was incorporated for interpretation of remedial data. The significance of the present strategy for constructing multivariant effects of electron donors could be objectively judged by pattern comparison with the short-term data analyzed. Thus, grafted aromatics as methyl- or ethylbenzene require much more microbial reaction time, even with mixed aromatic donors or stronger electron donors such as methanol in the original reduced medium, as indicated in the scatter chart. However, completion of the remedial time was needed by the kinetic simulation, and even low, smooth data were expressed. Among the exponential decay curves indicated, the carbon sources in the mix were favorably expressed. The smooth pattern indicated that fermentations with glucose and molasses showed lower remedial activity than melanoids or the indole series. The vigor increase was better for melanoid carbon in the initial fermentation of 24 h, while molasses increased later at 72 h and was more quenched by amending indole acetic acid (IAA) or indole expression. The molecular interaction of the electron mediator indole acetic acid in most trials indicated a quenched effect on toluene and ethylbenzene degradation, even when mixed with the original reducing medium, but expressed better with molasses in both kinetic simulations and growth effects. Thus, combining electron mediators such as IAA for Bacillus may offer a new degradation route for the metabolite alkylbenzene, which is worth further exploration for environmental aromatic waste remediation and combined restoration strategies.
Citation: HsiaoDao Chang, XiuYou Wan, HsiaoLan Huang, YiSu Chen, ChaoYing Chen. Anaerobic enrichment of Bacillus alkylbenzene remedial consortia from waste biomass melanoid sources[J]. AIMS Environmental Science, 2021, 8(4): 341-357. doi: 10.3934/environsci.2021022
Bioremediation of alkylbenzenes, including toluene, ethylbenzene and xylenes, was performed using fermentable aromatic sources and electron mediators by Bacillus cereus 301 in a limited oxygen state. The fermentation of small fermentable aromatic melanoids from cow manure as soluble humus hydrolysates or sugarcane molasses as saccharine, glucose and limited basal medium was compared. Thus, an evaluation model of exponential decline against a control was incorporated for interpretation of remedial data. The significance of the present strategy for constructing multivariant effects of electron donors could be objectively judged by pattern comparison with the short-term data analyzed. Thus, grafted aromatics as methyl- or ethylbenzene require much more microbial reaction time, even with mixed aromatic donors or stronger electron donors such as methanol in the original reduced medium, as indicated in the scatter chart. However, completion of the remedial time was needed by the kinetic simulation, and even low, smooth data were expressed. Among the exponential decay curves indicated, the carbon sources in the mix were favorably expressed. The smooth pattern indicated that fermentations with glucose and molasses showed lower remedial activity than melanoids or the indole series. The vigor increase was better for melanoid carbon in the initial fermentation of 24 h, while molasses increased later at 72 h and was more quenched by amending indole acetic acid (IAA) or indole expression. The molecular interaction of the electron mediator indole acetic acid in most trials indicated a quenched effect on toluene and ethylbenzene degradation, even when mixed with the original reducing medium, but expressed better with molasses in both kinetic simulations and growth effects. Thus, combining electron mediators such as IAA for Bacillus may offer a new degradation route for the metabolite alkylbenzene, which is worth further exploration for environmental aromatic waste remediation and combined restoration strategies.

| [1] |
Xiong F, Li Q, Zhou B et al. (2016) Oxidative stress and genotoxicity of long-term occupational exposure to low levels of BTEX in gas station workers. Int J Environ Res Pub Health 13: 1212. doi: 10.3390/ijerph13121212

|
| [2] |
Xing D, Magdouli S, Zhang J, et al. (2020) Microbial remediation for the removal of inorganic contaminants from treated wood: Recent trends and challenges. Chemosphere 258: 127429. doi: 10.1016/j.chemosphere.2020.127429
|
| [3] |
Baker DB, Ray PM (1965) Relation between effects of auxin on cell wall synthesis and cell elongation. Plant Physiol 41: 360-368. doi: 10.1104/pp.40.2.360
|
| [4] |
Janaki S, Thenmozhi S, Muthumari R (2016) A study on hydrocarbon degradation by biosurfactant producing Bacillus cereus in oil contaminated soil samples. Int J Life Sci Scienti Res 2: 324-332. doi: 10.21276/ijlssr.2016.2.4.4
|
| [5] |
Abbas SZ, Whui's TC, Hossain K, et al. (2018) Isolation and characterization of oil-degrading bacteria from marine sediment environment. Desalin Water Treat 136: 282-289. doi: 10.5004/dwt.2018.23110
|
| [6] | Ahmed IJ (2018) Biosurfactant and virulence factors are main tools of Bacillus cereus surviving. J Bacterial Mycol 6: 300-301. |
| [7] |
Berry DF, Francis AJ, Bollag JM et al. (1987) Microbial metabolism of homocyclic and heterocyclic aromatic compounds under anaerobic conditions. Microbiol Rev 51: 43-59. doi: 10.1128/mr.51.1.43-59.1987
|
| [8] | Ghosal D, Ghosh S, Dutta TK, et al. (2016) Current state of knowledge in microbial degradation of polycyclic aromatic hydrocarbons (PAHs): A Review. Front Microb 7: 1369. |
| [9] |
Spormann M, Widdel F (2000) Metabolism of alkylbenes, alkanes, and other hydrocarbons in anaerobic bacteria. Biodegradation11: 85-105. doi: 10.1023/A:1011122631799
|
| [10] |
Beller HR (2000) Metabolic indicators for detecting in situ anaerobic alkylbenezene degradation. Biodegradation11: 125-139. doi: 10.1023/A:1011109800916
|
| [11] | Das N, Chandran P (2011) Microbial degradation of petroleum hydrocarbon contaminants: An Overview. Biotech Res Int 2011: 941810. |
| [12] |
Feisthauer S, Seidel M, Bombach P, et al. (2012) Characterization of the relationship between microbial degradation processes at a hydrocarbon contaminated site using isotopic methods. J Contam Hydrol 133: 17-29. doi: 10.1016/j.jconhyd.2012.03.001
|
| [13] |
Nam K, Kukor JJ (2000) Combined ozone and biodegradation for remediation of mixtures of polycyclic aromatic hydrocarbons in soil. Biodegradation 11: 1-9. doi: 10.1023/A:1026592324693
|
| [14] | Galperin MY (2007) Some bacteria degrade explosives, others prefer boiling methanol. Environ Microbiol 12: 2905-2910. |
| [15] |
Grifoll M, Selifonov SA, Chapman P J, et al.(1994) Evidence for a novel pathway in the degradation of fluorene by Pseudomonas sp. strain F274. Appl Environ Microbiol 60: 2438-2449. doi: 10.1128/aem.60.7.2438-2449.1994
|
| [16] |
Yoshikawa M, Zhang M, Toyota K, et al. (2017) Biodegradation of volatile organic compounds and their effects on biodegradability under co-existing conditions. Microbes Environ 32: 188-200. doi: 10.1264/jsme2.ME16188
|
| [17] | Bashan Y, Puente ME, de-Bashan LE, et al. (2008) Environmental uses of plant growth-promoting bacteria. In: (Eds) E Ait Barka & C Clément, Plant-Microbe Interact 661: 69-73. |
| [18] |
Karthikeyan S, Radhakrishnan M (2010) Characterization of proton production and consumption associated with microbial metabolism. BMC Biotechnol 10: 2. doi: 10.1186/1472-6750-10-2
|
| [19] |
Lovley R (2000) Anaerobic benzene degradation. Biodegradation 11: 107-116. doi: 10.1023/A:1011191220463
|
| [20] |
Van Hamme JD, Singh A, Ward OP, et al. (2003) Recent advances in petroleum microbiology. Microbial Mol Bio Rev 67: 503-549. doi: 10.1128/MMBR.67.4.503-549.2003
|
| [21] |
Seo JS, Keum YS, Li QX, et al. (2009) Bacterial degradation of aromatic compounds. Int J Environ Res Public Health 6: 278-309. doi: 10.3390/ijerph6010278
|
| [22] | Phale PS and Shah BA 2020 Chapt. One-Degradation strategies and associated regulatory mechanisms/features for aromatic compound metabolism in bacteria. Adv Appl Micro 112: 1-65. |
| [23] |
Xin BP, Wu CH, Wu CH, et al. (2013) Bioaugmented remediation of high concentration BTEX-contaminated groundwater by permeable reactive barrier with immobilized bead. J Hazard Mater 244: 765-772. doi: 10.1016/j.jhazmat.2012.11.007
|
| [24] |
Lee EH (2009) Effect of substrate interaction the degradation of methyl tert-butyl ether, benzene toluene, ethylbenzene, and xylene by Rhodococcus sp. J Harzard Mater 167: 669-674. doi: 10.1016/j.jhazmat.2009.01.035
|
| [25] |
Kadri T, Magdouli A, Rouissi T, et al. (2018) Bench-scale production of enzymes from the hydrocarbonoclastic bacteria Alcanivoras borkumensis and biodegradation tests. J Biotechnol 283: 105-114. doi: 10.1016/j.jbiotec.2018.07.039
|
| [26] |
Kadri T, Rouissi T, Magdouli S, et al. (2018) Production and characterization of novel hydrocarbon degrading enzymes from Alcanivorax borkumensis. Int J Biol Marcromol 112: 230-240. doi: 10.1016/j.ijbiomac.2018.01.177
|
| [27] |
Kadri T, Magdouli S, Rouissi T, et al. (2018) Ex-situ biodegradation of petroleum hydrocarbons using Alcanivorax borkumensis enzymes. Biochem Eng J 132: 279-287. doi: 10.1016/j.bej.2018.01.014
|
| [28] | Adzitey F, Assoah-Peprah P, Teye AG, et al. (2020) Prevalence and antimicrobial resistance of Escherichia coli isolated from various meat types in the Tamale Metropolis of Ghana. Int J Food Sci 2020 |
| [29] |
Zigha A, Rosenfeld E, Schmitt P, et al. (2006) Anaerobic cells of Bacillus cereus F4430/73 respond to low oxidoreduction potential by metabolic readjustments and activation of enterotoxin expression. Arch Microbiol 185: 222-233. doi: 10.1007/s00203-006-0090-z
|
| [30] | Rosenfeld E, Duport C, Zigha A, et al. (2005) Characterization of aerobic and anaerobic vegetative growth of the food-borne pathogen Bacillus cereus F4430/73 strain. Can J Microbiol 51 : 149-158. |
| [31] | Cason ED, Vermeulen JG, Muller WJ, et al. (2019) Aerobic and anaerobic enrichment cultures highlight the pivotal role of facultative anaerobes in soil hydrocarbon degradation. J Environ Sci Heal Par A Tox /Haz Sub & Environ Eng 54: 408-415. |
| [32] |
Dou JF, Wang JS, Liu XA et al. (2010) Anaerobic benzene biodegradation by a pure bacterial culture of Bacillus cereus under nitrate reducing conditions. J Environ Sci 22: 709-715. doi: 10.1016/S1001-0742(09)60167-4
|
| [33] |
Masciarelli O, Llanes A, Luna V (2014) A new PGPR co-inoculated with Bradyrhizobium japonicumenhances soybean nodulation. Microbiol Res169: 609-615. doi: 10.1016/j.micres.2013.10.001
|
| [34] | Zhaoa YT, Songa CI, Dong HQ, et al. (2018) Community structure and distribution of culturable bacteria in soil along an altitudinal gradient of Tianshan Mountains, China Biotech & Biotechnol Eq 32: 397-407. |
| [35] |
Zhang X, Sullivan ER, Young LY, et al. (2000) Evidence for aromatic ring reduction in the biodegradation pathway of carboxylate naphthalene by a sulfate reducing consortium. Biodegradation 11: 117-124. doi: 10.1023/A:1011128109670
|
| [36] |
Nikitin N (2018) Impact of effective microorganisms on the transfer of radioactive cesium into lettuce and barley biomass. J Environ Rad 192: 491-497. doi: 10.1016/j.jenvrad.2018.08.005
|
| [37] |
Osborn RK, Haydock PPJ, Edwards SG (2010) Isolation and identification of oxamyl-degrading bacteria from UK agricultural soils. Soil Biol Biochem 42: 998-1000. doi: 10.1016/j.soilbio.2010.01.016
|
| [38] |
Vogt C, Kleinsteuber S, Richnow HH (2011) Anaerobic benzene degradation by bacteria. Microb Biotechnol 4: 710-724. doi: 10.1111/j.1751-7915.2011.00260.x
|
| [39] |
Caldwell ME and Suflita JM (2000) Detection of phenol and benzoate as intermediates of anaerobic benzene biodegradation under different terminal electron-accepting conditions. Environ Sci Tech 34: 1216-1220. doi: 10.1021/es990849j
|
| [40] |
van der Waals MJ, Siavash A, da Rocha UN, et al. (2017) Degradation in a denitrifying biofilm reactor: activity and microbial community composition. Appl Microbiol Biotechnol 101: 5175-5188. doi: 10.1007/s00253-017-8214-8
|
| [41] |
Patowary K, Patowary R, Kalita MC, et al. (2016) Development of an efficient bacterial consortium for the potential remediation of hydrocarbons from contaminated sites. Front Microbiol 7:1092. doi: 10.3389/fmicb.2016.01092
|
| [42] |
Kamal MA, Klein P (2010) Estimation of BTEX in groundwater by using gas chromatography-mass spectrometry. Saudi J Biol Sci 17: 205-208. doi: 10.1016/j.sjbs.2010.04.002
|
| [43] |
Zhao L, QinX, Hou X, et al. (2018) Research on determination of BTEX in human whole blood using purge and trap-gas chromatography-mass spectrometry combined with isotope internal standard. Microchemical J 145: 308-312. doi: 10.1016/j.microc.2018.10.038
|
| [44] |
Cook RL, Hesterberg D (2013) Comparison of trees and grasses for rhizome mediation of petroleum hydrocarbons. Int J Phytoremediation 15: 844-860. doi: 10.1080/15226514.2012.760518
|
| [45] |
Xu WL, Wu LL, Ma L, et al.(2015) Effects of Indole-3-acetic acid (IAA), a plant hormone, on the ryegrass yield and the removal of fluoranthene from soil. Intl J Phytoremediation 17: 422-428. doi: 10.1080/15226514.2014.910172
|
| [46] | El-Shafei A, Mahmoud Y, El-Naqib F, et al. (2008) Impact of effective microorganism compost on soil fertility and rice productivity and quality. Misr J Ag Eng 25:1067-1093. |
| [47] |
Highton PJ, Hobbs DG (1971) Penicillin and cell wall synthesis: a study of Bacillus licheniformis by electron microscopy. J Bacterio l 106: 646-658. doi: 10.1128/jb.106.2.646-658.1971
|
| [48] |
Meléndez-Hevia E, Waddell TG, Cascante M (1996)The puzzle of the Krebs citric acid cycle: assembling the pieces of chemically feasible reactions, and opportunism in the design of metabolic pathways during evolution. J Mol Evol 43: 293-303. doi: 10.1007/BF02338838
|
| [49] | Mohite B (2013) Isolation and characterization of indole acetic acid (IAA) producing bacteria from rhizospheric soil and its effect on plant growth. J Soil Sci Plant Nutr 13: 638-649. |
| [50] |
Arkhipchenko IA, Shaposhnikov AI, Kravchenko LV, et al. (2006) Tryptophan concentration of animal wastes and organic fertilizers. Appl Soil Ecol 34: 62-64. doi: 10.1016/j.apsoil.2005.12.008
|
| [51] |
Bianco C, Imperlini E, Calogero R, et al. (2006) Indole-3-acetic acid improves Escherichia coli's defences to stress. Arch Microbiol 185: 373-82. doi: 10.1007/s00203-006-0103-y
|
| [52] | Wahyudi T, Astuti RP, Widyawati A, et al. (2011) Characterization of Bacillus sp. strains isolated from rhizosphere of soybean plants for their use as potential plant growth for promoting rhizobacteria. J Microb Antimicrob 3: 34-40. |
Figures(6) / Tables(2)

HsiaoDao Chang, XiuYou Wan, HsiaoLan Huang, YiSu Chen, ChaoYing Chen. Anaerobic enrichment of Bacillus alkylbenzene remedial consortia from waste biomass melanoid sources[J]. AIMS Environmental Science, 2021, 8(4): 341-357. doi: 10.3934/environsci.2021022







 DownLoad:
DownLoad: