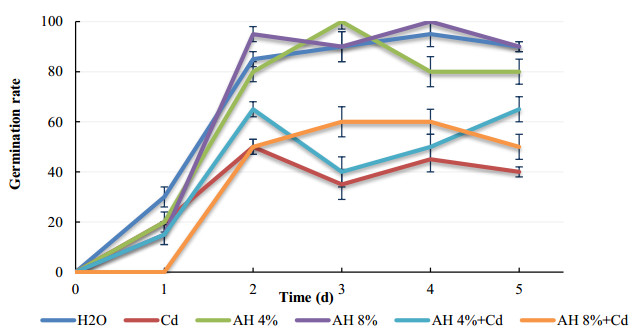
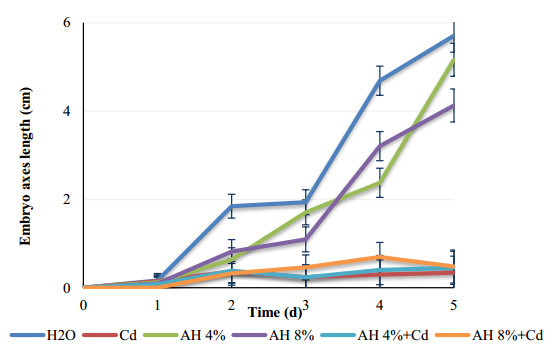
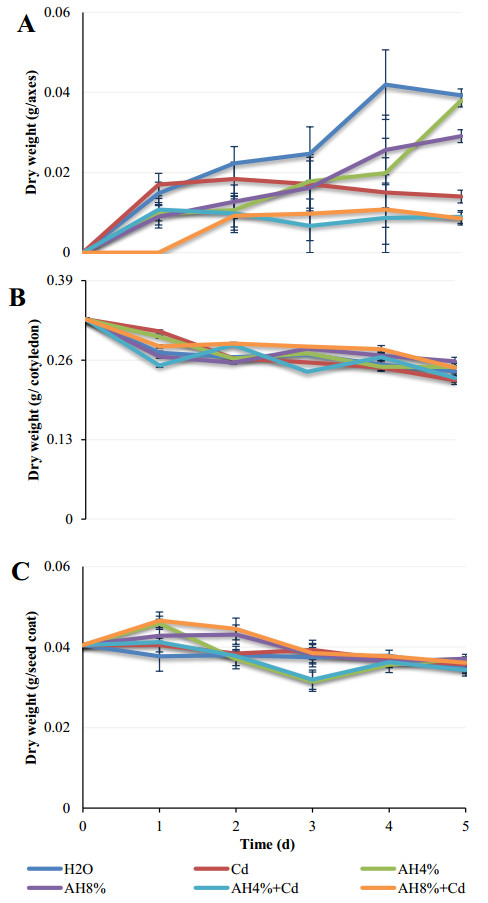
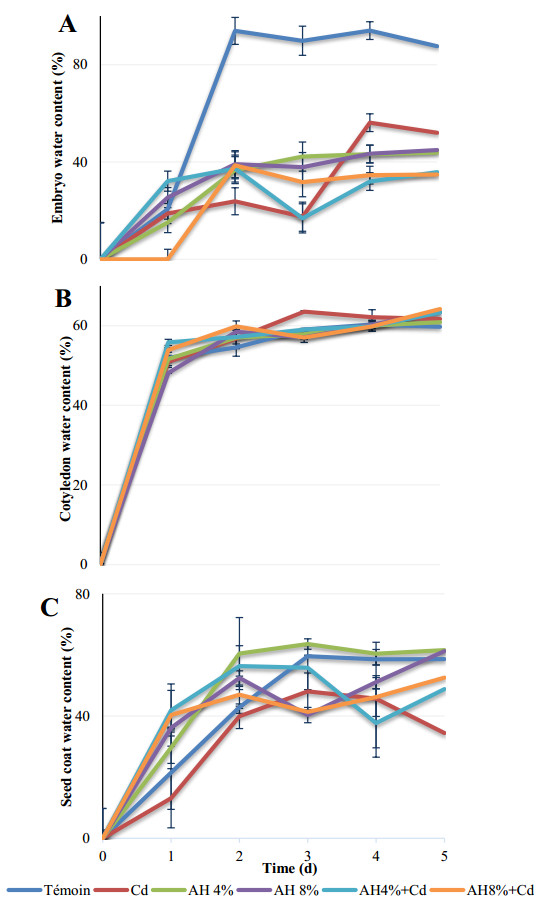
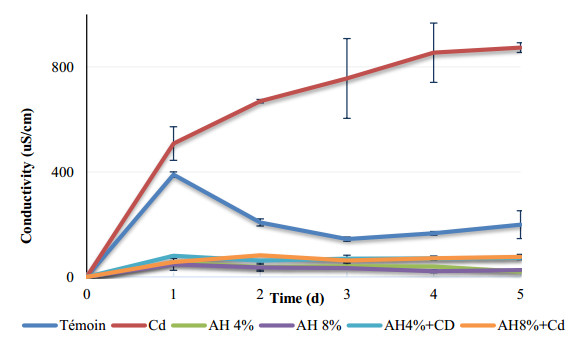
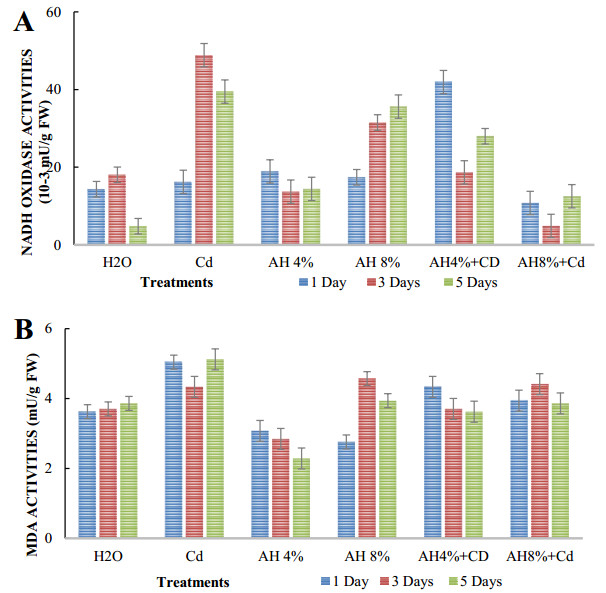
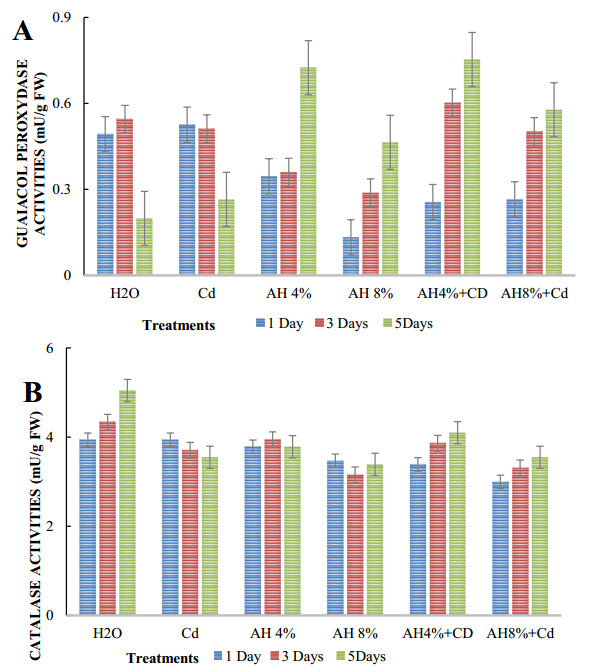
Seeds of pea (Pisum sativum L. var. Alicia) were germinated for 5 d by soaking in distilled water or treated with cadmium and/or humic acid (HA). Interaction between humic acid and cadmium was previously discussed. However, the regulation of the humic acid / cadmium interaction during seed germination, the first crucial stage of the plant life cycle, need to be clarified. Seed germination is an important stage of the plant life, which is highly sensitive to surrounding medium changes, since the germinating seed is the first interface of material exchange between plant cycle and environment. Relationships among cadmium stress, HA-treatment, germination rate, and changes in reserve mobilization were studied. Cadmium disrupted the soaking process that was a major event of germination. Moreover, important solutes leakage due to Cd-treatment caused disorder in reserve mobilization from cotyledons to growing embryonic axes. The observed delay in germination rate and seedling growth after Cd-treatment could be explained, partially, by the disturbance in the mobilization of water and nutrients. NADH-and MDA-activities were considered as markers of oxidative stress. they were substantially increased after Cd-treatment compared with controls but decreased in presence of AH. Excessive NADH oxidase activities proved the install of oxidative stress in Cd poisoned seeds. We suggested an antagonist effect of Cd and HA on oxidative stress enzyme activities. Treatment of seeds with Cd and/or AH maintained a high level of guaiacol peroxidase activities during five days. The possible implication of GPOX against Cd toxicity and the improvement of the antioxidant system after AH treatment proved an important implication of these findings during germination.
Citation: Mohammed Basahi. Humic acid improved germination rate, seedling growth and antioxidant system of pea (Pisum sativum L. var. Alicia) grown in water polluted with CdCl2[J]. AIMS Environmental Science, 2021, 8(4): 358-370. doi: 10.3934/environsci.2021023
Seeds of pea (Pisum sativum L. var. Alicia) were germinated for 5 d by soaking in distilled water or treated with cadmium and/or humic acid (HA). Interaction between humic acid and cadmium was previously discussed. However, the regulation of the humic acid / cadmium interaction during seed germination, the first crucial stage of the plant life cycle, need to be clarified. Seed germination is an important stage of the plant life, which is highly sensitive to surrounding medium changes, since the germinating seed is the first interface of material exchange between plant cycle and environment. Relationships among cadmium stress, HA-treatment, germination rate, and changes in reserve mobilization were studied. Cadmium disrupted the soaking process that was a major event of germination. Moreover, important solutes leakage due to Cd-treatment caused disorder in reserve mobilization from cotyledons to growing embryonic axes. The observed delay in germination rate and seedling growth after Cd-treatment could be explained, partially, by the disturbance in the mobilization of water and nutrients. NADH-and MDA-activities were considered as markers of oxidative stress. they were substantially increased after Cd-treatment compared with controls but decreased in presence of AH. Excessive NADH oxidase activities proved the install of oxidative stress in Cd poisoned seeds. We suggested an antagonist effect of Cd and HA on oxidative stress enzyme activities. Treatment of seeds with Cd and/or AH maintained a high level of guaiacol peroxidase activities during five days. The possible implication of GPOX against Cd toxicity and the improvement of the antioxidant system after AH treatment proved an important implication of these findings during germination.

| [1] | Smiri M, Bousami S, Missaoui T, et al. (2016) The cadmium-binding thioredoxin o acts as an upstream regulator of the redox plant homeostasis. In Redox State as a Central Regulator of Plant-Cell Stress Responses. Edited by Gupta DK, Palma JM and Corpas FJ. Springer International Publishing Switzerland, 350. |
| [2] |
Smiri M, Missaoui T (2014) The role of ferredoxin: thiorédoxine reductase/ thioredoxin m in seed germination and the connection between this system and copper ion toxicity. J Plant Physiol 171: 1664-1670. doi: 10.1016/j.jplph.2014.06.019

|
| [3] | Missaoui T, Smiri M, Chemingui H, et al (2021) Effect of Nanosized TiO2 on Redox Properties in Fenugreek (Trigonella foenum graecum L.) during Germination. Environ Process 8: 843-867. |
| [4] | Juste C, Chassin P, Gomez A, et al. (1995) Les micro-polluants métalliques dans les boues résiduaires des stations d'épuration urbaines. Convention Ademe. |
| [5] | Kabata-Pendias A, Pendias H (1992) Trace elements in soils and plants. Boca Raton: CRC Press 365. |
| [6] | Chemingui H, Smiri M, Missaoui T, et al. (2019) Zinc Oxide Nanoparticles Induced Oxidative Stress and Changes in the Photosynthetic Apparatus in Fenugreek (Trigonella foenum graecum L.). Bull Environ Cont Toxicol 102: 477-485. |
| [7] |
Li Y, Fang F, Wei J, et al. (2019) Humic Acid Fertilizer Improved Soil Properties and Soil Microbial Diversity of Continuous Cropping Peanut: A Three-Year Experiment. Sci Rep 9: 12014. doi: 10.1038/s41598-019-48620-4
|
| [8] |
Xu M, Zhao Z, Shi M, et al. (2019) Effect of humic acid on the stabilization of cadmium in soil by coprecipitating with ferrihydrite. Environ Sci Pollut Res 26: 27330-27337. doi: 10.1007/s11356-019-05893-6
|
| [9] | Karlsson T (2005) Complexation of cadmium, copper and methyl mercury to functional groups in natural organic matter. Thèse, Faculty of Forest Sciences, Umea, Suède, 37. |
| [10] | Smiri M, Chaoui A, Rouhier N, et al. (2010) NAD pattern and NADH oxidase activity in pea (Pisum sativum L.) under cadmium toxicity. Physiology and Molecular Biology of Plants 16: 305-315. |
| [11] |
Missaoui T, Smiri M, Hafiane A (2019) Reserve Mobilization, Membrane Damage and Solutes Leakage in Fenugreek Imbibed with Urban Treated Wastewater. Bull Environ Cont Toxicol 103: 461-467. doi: 10.1007/s00128-019-02658-2
|
| [12] | Smiri M (2011) Effect of cadmium on germination, growth, redox and oxidative properties in Pisum sativum seeds. Journal of Environmental Chemistry and Ecotoxicology 3: 52-59. |
| [13] | Smiri M, Jelali N, El Ghoul J (2013) Cadmium affects the NADP-thioredoxinreductase/thioredoxin system in germinating pea seeds. J Plant Inter 8: 125-133. |
| [14] | Hartwigsen JA, Evans MR (2000) Humic acids seed and substrate treatments promote seedling root development. Hort Science 35: 1231-1233. |
| [15] |
Smiri M, Elarbaoui S, Missaoui T, et al. (2015) Micropollutants in Sewage Sludge: Elemental Composition and Heavy Metals Uptake by Phaseolus vulgaris and Vicia faba Seedlings. Arab J Sci Eng 40: 1837-1847. doi: 10.1007/s13369-015-1639-4
|
| [16] |
Witcombe JR, Hollington PA, Howarth CJ, et al. (2009) Breeding for abiotic stresses for sustainable agriculture. Phil Trans R Soc B 363: 703-716. doi: 10.1098/rstb.2007.2179
|
| [17] | Bouzerzour H, Benmahammed A (2009) Variation in early growth, canopy temperature, translocation and yield of four durum wheat (Triticum durum Desf.) genotypes under semi-arid conditions. J Agric Sci 5: 142-154. |
| [18] | Hussain SS (2006) Molecular breeding for abiotic stress tolerance: Drought perspective. Proc Pak Acad Sci 43:189-210. |
| [19] | Vassilev A, Yordanov I (1997) Reductive analysis of factors limiting growth of cadmium-treated plants: a review. Bulgarian J Plant Physiol 23: 114-133. |
| [20] | Chen Y, Aviad T (1990) Effects of Humic Substance on Plant Growth.In Humic Substances in soil and Crop Sciences. Soil Sci Soc America 1990: 161-186. |
| [21] |
De Kreij C (1995) Effect of humic substances in nutrient film technique on nutrient uptake. J Plant Nut 18: 793-802. doi: 10.1080/01904169509364938
|
| [22] |
Rauthan BS, Schnitzer M (1981) Effects of a soil fulvic acid on the growth and nutrient content of cucumber (Cucumis sativus) plants. Plant Soil 63: 491-495. doi: 10.1007/BF02370049
|
| [23] | Bauduin M, Delcarte E, Impens R (1986) Impact des composts urbains sur les teneurs en Zn et en Cu des tissus végétaux. Deuxième Symposium International sur le rôle des ligoéléments en Agriculture, Toulouse, 441-449. |
| [24] | Fernandez-Escobar R, Benlloch M, Barranco D, et al. (1996) Response of olive trees to foliar application of humic substances extracted from leonardite. Sci Hort 66: 191-200. |
| [25] |
Fagbenro JA, Agboola AA (1993) Effect of different levels of humic acid on the growth and nutrient uptake of teak seedlings. J Plant Nutr 16: 1465-1483. doi: 10.1080/01904169309364627
|
| [26] |
Ayuso M, Hernández T, Garcia C, et al. (1996) Stimulation of barley growth and nutrient absorption by humic substances originating from various organic materials. Biores Tech 57: 251-257. doi: 10.1016/S0960-8524(96)00064-8
|
| [27] |
Canellas LP, Olivares FL, Okorokovha-Façanha AI, et al. (2002) Humic acids isolated from earthworm compost enhance root elongation, lateral root emergence and plasma membrane H+-ATPase activity in maize root. Plant Physiol 130: 1951-1957. doi: 10.1104/pp.007088
|
| [28] |
Koca H, Bor M, Özdemir F, et al. (2007) The effect of salt stress on lipid peroxidation, antioxidative enzymes and proline content of sesame cultivars. Environ Exp Bot 60: 344-351. doi: 10.1016/j.envexpbot.2006.12.005
|
| [29] | Gill SS, Khan NA, Tuteja N (2012) Cadmium at high dose perturbs growth, photosynthesis and nitrogen metabolism while at low dose it up regulates sulphur assimilation and antioxidant machinery in garden cress (Lepidiumsativum L.). Plant Sci 182: 112-120. |
| [30] | Moller IM (2001) Plant mitochondria and oxidative stress: Electron transport, NADPH turnover, and metabolism of reactive oxygen species. Ann. Rev. Plant Physiol. Plant Mol Biol 52: 561-591. |
| [31] | Singh S, Khan NA, Nazar R, et al. (2008) Photosynthetic traits and activities of antioxidant enzymes in blackgram (Vignamungo L. Hepper) under cadmium stress. Am J Plant Physiol 3: 25-32. |
| [32] | Metwally A, Safronova VI, Belimov AA, et al. (2005) Genotypic variation of the response to cadmium toxicity in Pisum sativum L. J Exp Bot 56: 167-178. |
| [33] |
Ahsan N, Lee SH, Lee DG, et al. (2007) Physiological and protein profiles alternation of germinating rice seedlings exposed to acute cadmium toxicity. C R Biol 330: 735-746. doi: 10.1016/j.crvi.2007.08.001
|
| [34] |
Filek M, Keskinen R, Hartikainen H, et al. (2008) The protective role of selenium in rape seedlings subjected to cadmium stress. J Plant Physiol 165: 833-844. doi: 10.1016/j.jplph.2007.06.006
|
| [35] |
Romero-Puertas MC, Rodriguez-Serrano M, Corpas FJ, et al. (2004) Cadmium induced subcellular accumulation of O2- and H2O2 in pealeaves. Plant Cell Environ 27: 1122-1134. doi: 10.1111/j.1365-3040.2004.01217.x
|
| [36] |
Mobin M, Khan NA (2007) Photosynthetic activity, pigment composition and antioxidative response of two mustard (Brassica juncea) cultivars differing in photosynthetic capacity subjected to cadmium stress. J Plant Physiol 164: 601-610. doi: 10.1016/j.jplph.2006.03.003
|
| [37] |
Balestrasse KB, Noriega GO, Batlle A, et al. (2006) Heme oxygenase activity and oxidative stress signaling in soybean leaves. Plant Sci 170: 339-346. doi: 10.1016/j.plantsci.2005.09.001
|
| [38] |
Guo TR, Zhang GP, Zhou MX, et al. (2007) Influence of aluminum andcadmium stresses on mineral nutrition and rootexudates in two barley cultivars. Pedosphere 17: 505-512. doi: 10.1016/S1002-0160(07)60060-5
|
| [39] |
Nardi S, Pizzeghello D, Muscolo A, et al. (2002) Physiological effects of humic substances on higher plants. Soil Biol Biochem 34: 1527-1536. doi: 10.1016/S0038-0717(02)00174-8
|
| [40] |
Bradl HB (2004) Adsorption of heavy metal ions on soils and soils constituents. J Coll Inter Sci 277: 1-18. doi: 10.1016/j.jcis.2004.04.005
|
| [41] | Deschamps T, Bansaazoua M, Bussière B, et al. (2006) Mécanismes de rétention des métaux lourds en phase solide: Cas de la stabilisation des sols contaminés et des déchets industriels. VertigO, La revue électronique en sciences de l'environnement, 7: 2. |
| [42] |
Hettiarachchi GM, Ryan JA, Chaney RL, et al. (2003) Sorption and desorption of cadmium by different fractions of biosolids-amended soils. J Environ Quality 32: 1684-1693. doi: 10.2134/jeq2003.1684
|
Figures(7)

Mohammed Basahi. Humic acid improved germination rate, seedling growth and antioxidant system of pea (Pisum sativum L. var. Alicia) grown in water polluted with CdCl2[J]. AIMS Environmental Science, 2021, 8(4): 358-370. doi: 10.3934/environsci.2021023







 DownLoad:
DownLoad: