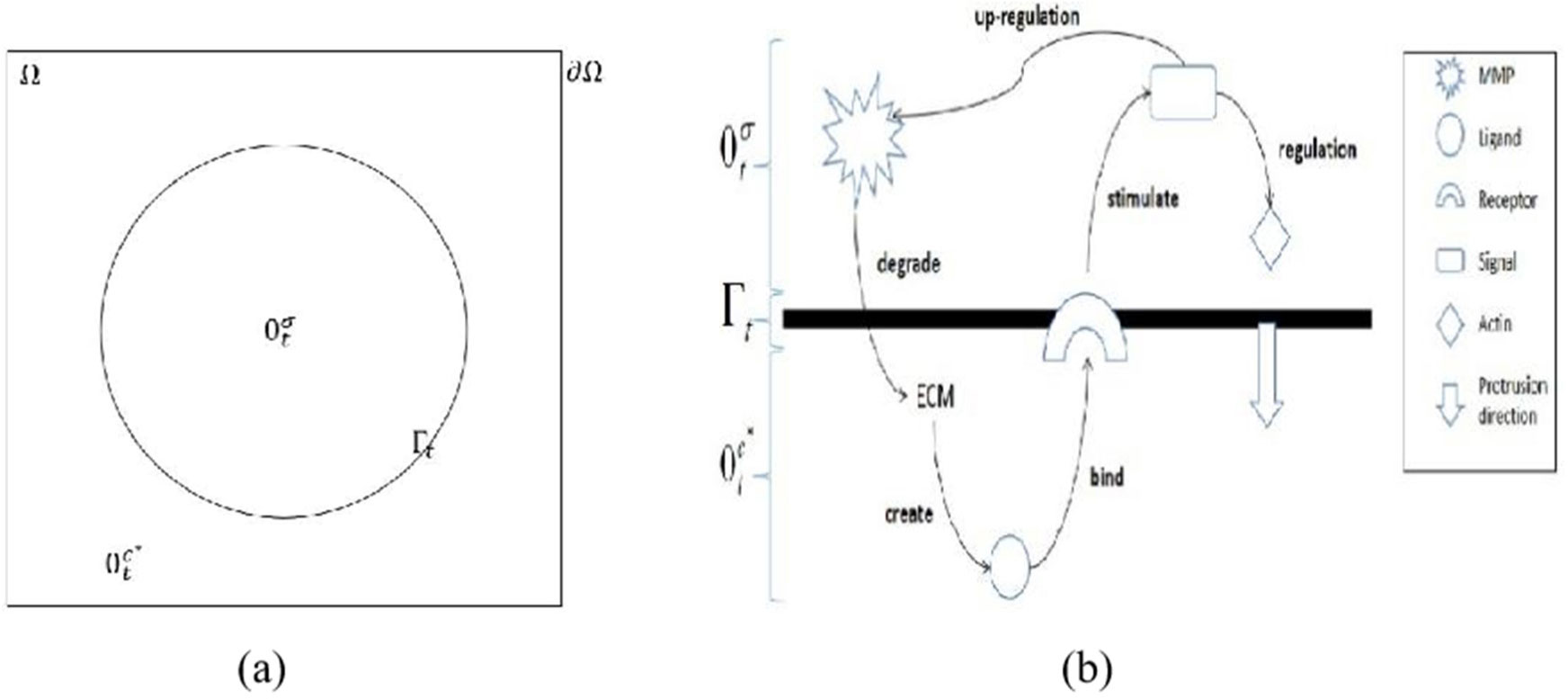
Invadopodia are finger-like protrusions that are commonly spotted at the membrane of the invasive cancer cell. These structures are a major cause of death among cancer patients through metastasis process. Signal transduction stimulated upon contact between ligand and membrane receptors is identified as one of key factors in invadopodia formation. In this study, a time-dependent mathematical model of signal and ligand is investigated numerically. The moving boundary of plasma membrane is taken as a zero-level set function and is moved by the velocity that accounted as the difference of gradient between intra-cellular signal and extra-cellular ligand. The model is solved using a combination of ghost with linear extrapolation and finite difference methods. The results showed that the stimulation of signal from membrane associated ligand consequently moved the plasma membrane outward as time increases. The highest densities of signal and ligand are recorded on the membrane and slowly diffused into intra-cellular and extra-cellular regions, respectively.
Citation: Noorehan Yaacob, Sharidan Shafie, Takashi Suzuki, Mohd Ariff Admon. Signal transduction from ligand-receptor binding associated with the formation of invadopodia in an invasive cancer cell[J]. AIMS Bioengineering, 2022, 9(3): 252-265. doi: 10.3934/bioeng.2022017
Invadopodia are finger-like protrusions that are commonly spotted at the membrane of the invasive cancer cell. These structures are a major cause of death among cancer patients through metastasis process. Signal transduction stimulated upon contact between ligand and membrane receptors is identified as one of key factors in invadopodia formation. In this study, a time-dependent mathematical model of signal and ligand is investigated numerically. The moving boundary of plasma membrane is taken as a zero-level set function and is moved by the velocity that accounted as the difference of gradient between intra-cellular signal and extra-cellular ligand. The model is solved using a combination of ghost with linear extrapolation and finite difference methods. The results showed that the stimulation of signal from membrane associated ligand consequently moved the plasma membrane outward as time increases. The highest densities of signal and ligand are recorded on the membrane and slowly diffused into intra-cellular and extra-cellular regions, respectively.

| [1] |
Helfinger V, Schröder K (2018) Redox control in cancer development and progression. Mol Aspects Med 63: 88-98. https://doi.org/10.1016/j.mam.2018.02.003 
|
| [2] |
Li B, Gordon GM, Du CH, et al. (2010) Specific killing of Rb mutant cancer cells by inactivating TSC2. Cancer Cell 17: 469-480. https://doi.org/10.1016/j.ccr.2010.03.019
|
| [3] |
Szabó A, Merks RMH (2013) Cellular potts modeling of tumor growth, tumor invasion, and tumor evolution. Front Oncol 3: 87. https://doi.org/10.3389/fonc.2013.00087
|
| [4] |
Franssen LC, Lorenzi T, Burgess AEF, et al. (2019) A mathematical framework for modelling the metastatic spread of cancer. Bull Math Biol 81: 1965-2010. https://doi.org/10.1007/s11538-019-00597-x
|
| [5] |
Jiang WG, Sanders AJ, Katoh M, et al. (2015) Tissue invasion and metastasis: Molecular, biological and clinical perspectives. Semin Cancer Biol 35: S244-S275. https://doi.org/10.1016/j.semcancer.2015.03.008
|
| [6] |
Meirson T, Gil-Henn H (2018) Targeting invadopodia for blocking breast cancer metastasis. Drug Resist Update 39: 1-17. https://doi.org/10.1016/j.drup.2018.05.002
|
| [7] |
Sibony-Benyamini H, Gil-Henn H (2012) Invadopodia: the leading force. Eur J Cell Biol 91: 896-901. https://doi.org/10.1016/j.ejcb.2012.04.001
|
| [8] |
Parekh A, Weaver AM (2016) Regulation of invadopodia by mechanical signaling. Exp Cell Res 343: 89-95. https://doi.org/10.1016/j.yexcr.2015.10.038
|
| [9] |
Deakin NE, Chaplain MAJ (2013) Mathematical modeling of cancer invasion: The role of membrane-bound matrix metalloproteinases. Front Oncol 3: 70. https://doi.org/10.3389/fonc.2013.00070
|
| [10] |
Chaplain M, Lolas (2006) Mathematical modelling of cancer invasion of tissue: dynamic heterogeneity. Networks Heterog Media 1: 399-439. https://doi.org/10.3934/nhm.2006.1.399
|
| [11] |
Ramis-Conde I, Chaplain MAJ, Anderson ARA (2008) Mathematical modelling of cancer cell invasion of tissue. Math Comput Model 47: 533-545. https://doi.org/10.1016/j.mcm.2007.02.034
|
| [12] |
Sfakianakis N, Madzvamuse A, Chaplain MAJ (2020) A hybrid multiscale model for cancer invasion of the extracellular matrix. Multiscale Model Simul 18: 824-850. https://doi.org/10.1137/18M1189026
|
| [13] |
Nska ZS, Rodrigo CM, Lachowicz M, et al. (2009) Tissue: the role and effect of nonlocal. Math Model Methods Appl Sci 19: 257-281. https://doi.org/10.1142/S0218202509003425
|
| [14] |
Saitou T, Rouzimaimaiti M, Koshikawa N, et al. (2012) Mathematical modeling of invadopodia formation. J Theor Biol 298: 138-146. https://doi.org/10.1016/j.jtbi.2011.12.018
|
| [15] | Admon MA (2015) Mathematical modeling and simulation in an individual cancer cell associated with invadopodia formation [PhD Thesis]. Japan: Osaka University. |
| [16] |
Gallinato O, Ohta M, Poignard C, et al. (2017) Free boundary problem for cell protrusion formations: theoretical and numerical aspects. J Math Biol 75: 263-307. https://doi.org/10.1007/s00285-016-1080-7
|
| [17] |
Admon MA, Suzuki T (2017) Signal transduction in the invadopodia formation using fixed domain method. J Phys Conf Ser 890: 012036. https://doi.org/10.1088/1742-6596/890/1/012036
|
| [18] | Azhuan NAN, Poignard C, Suzuki T, et al. (2019) Two-dimensional signal transduction during the formation of invadopodia. Malaysian J Math Sci 13: 155-164. |
| [19] |
Gallinato O, Colin T, Saut O, et al. (2017) Tumor growth model of ductal carcinoma: from in situ phase to stroma invasion. J Theor Biol 429: 253-266. https://doi.org/10.1016/j.jtbi.2017.06.022
|
| [20] |
Gallinato O, Poignard C (2017) Superconvergent second order Cartesian method for solving free boundary problem for invadopodia formation. J Comput Phys 339: 412-431. https://doi.org/10.1016/j.jcp.2017.03.010
|
| [21] |
Yaacob N, Noor Azhuan NA, Shafie S, et al. (2019) Numerical computation of signal stimulation from ligand-EGFR binding during invadopodia formation. Matematika 35: 139-148. https://doi.org/10.11113/matematika.v35.n4.1268
|
| [22] |
Osher S, Sethian JA (1988) Fronts propagating with curvature-dependent speed: Algorithms based on Hamilton-Jacobi formulations. J Comput Phys 79: 12-49. https://doi.org/10.1016/0021-9991(88)90002-2
|
| [23] |
Sethian JA (1996) Theory, algorithms, and applications of level set methods for propagating interfaces. Acta Numer 5: 309-395. https://doi.org/10.1017/S0962492900002671
|
| [24] |
Chen S, Merriman B, Osher S, et al. (1997) A simple level set method for solving stefan problems. J Comput Phys 135: 8-29. https://doi.org/10.1006/jcph.1997.5721
|
| [25] |
Tan L, Zabaras N (2006) A level set simulation of dendritic solidification with combined features of front-tracking and fixed-domain methods. J Comput Phys 211: 36-63. https://doi.org/10.1016/j.jcp.2005.05.013
|
| [26] |
Tan L, Zabaras N (2007) A level set simulation of dendritic solidification of multi-component alloys. J Comput Phys 221: 9-40. https://doi.org/10.1016/j.jcp.2006.06.003
|
| [27] |
Tan L, Zabaras N (2007) Modeling the growth and interaction of multiple dendrites in solidification using a level set method. J Comput Phys 226: 131-55. https://doi.org/10.1016/j.jcp.2007.03.023
|
| [28] |
Sethian JA, Adalsteinsson D (1999) The fast construction of extension velocities in level set methods. J Comput Phys 148: 2-22. https://doi.org/10.1006/jcph.1998.6090
|
| [29] |
Shashni B, Ariyasu S, Takeda R, et al. (2018) Size-based differentiation of cancer and normal cells by a particle size analyzer assisted by a cell-recognition PC software. Biol Pharm Bull 41: 487-503. https://doi.org/10.1248/bpb.b17-00776
|
| [30] |
Li A, Dawson JC, Forero-Vargas M, et al. (2010) The actin-bundling protein fascin stabilizes actin in invadopodia and potentiates protrusive invasion. Curr Biol 20: 339-345. https://doi.org/10.1016/j.cub.2009.12.035
|
| [31] |
Oser M, Mader CC, Gil-Henn H, et al. (2010) Specific tyrosine phosphorylation sites on cortactin regulate Nck1-dependent actin polymerization in invadopodia. J Cell Sci 123: 3662-3673. https://doi.org/10.1242/jcs.068163
|
| [32] |
Ren XL, Qiao YD, Li JY, et al. (2018) Cortactin recruits FMNL2 to promote actin polymerization and endosome motility in invadopodia formation. Cancer Lett 419: 245-256. https://doi.org/10.1016/j.canlet.2018.01.023
|
| [33] |
Drummond ML, Li M, Tarapore E, et al. (2018) Actin polymerization controls cilia-mediated signaling. J Cell Biol 217: 3255-3266. https://doi.org/10.1083/jcb.201703196
|
| [34] |
Singh AB, Harris RC (2005) Autocrine, paracrine and juxtacrine signaling by EGFR ligands. Cell Signal 17: 1183-1193. https://doi.org/10.1016/j.cellsig.2005.03.026
|
| [35] |
Walker D, Wood S, Southgate J, et al. (2006) An integrated agent-mathematical model of the effect of intercellular signalling via the epidermal growth factor receptor on cell proliferation. J Theor Biol 242: 774-789. https://doi.org/10.1016/j.jtbi.2006.04.020
|
| [36] |
Alroy I, Yarden Y (1997) The ErbB signaling network in embryogenesis and oncogenesis: Signal diversification through combinatorial ligand-receptor interactions. FEBS Lett 410: 83-86. https://doi.org/10.1016/S0014-5793(97)00412-2
|
| [37] |
Higashiyama S, Iwabuki H, Morimoto C, et al. (2008) Membrane-anchored growth factors, the epidermal growth factor family: beyond receptor ligands. Cancer Sci 99: 214-220. https://doi.org/10.1111/j.1349-7006.2007.00676.x
|
| [38] | Hoshino D, Branch KM, Weaver AM (2013) Signaling inputs to invadopodia and podosomes. J Cell Sci 126: 2979-89. https://doi.org/10.1242/jcs.079475 |
 bioeng-09-03-017-s001.pdf bioeng-09-03-017-s001.pdf |
 |




Figures(5) / Tables(2)

Noorehan Yaacob, Sharidan Shafie, Takashi Suzuki, Mohd Ariff Admon. Signal transduction from ligand-receptor binding associated with the formation of invadopodia in an invasive cancer cell[J]. AIMS Bioengineering, 2022, 9(3): 252-265. doi: 10.3934/bioeng.2022017







 DownLoad:
DownLoad: