Citation: Matthew Hogan, Glauco Souza, Ravi Birla. Assembly of a functional 3D primary cardiac construct using magnetic levitation[J]. AIMS Bioengineering, 2016, 3(3): 277-288. doi: 10.3934/bioeng.2016.3.277

| [1] |
Ehler E, Jayasinghe SN (2014) Cell electrospinning cardiac patches for tissue engineering the heart. Analyst 139: 4449–52. doi: 10.1039/C4AN00766B

|
| [2] | Haraguchi Y, Shimizu T, Matsuura K, et al. (2014) Cell sheet technology for cardiac tissue engineering. Methods Mol Biol 1181: 139c55. |
| [3] | Hirt MN, Hansen A, Eschenhagen T (2014) Cardiac tissue engineering: state of the art. Circ Res 114: 354–67. |
| [4] | Macadangdang J, Lee HJ, Carson D, et al. (2014) Capillary force lithography for cardiac tissue engineering. J Vis Exp (88): 50039. |
| [5] | Matsuura K, Masuda S, Shimizu T (2014) Cell sheet-based cardiac tissue engineering. AnatRec(Hoboken) 297: 65–72. |
| [6] |
Rao C, Barratt H, Prodromakis T, et al. (2014) Tissue engineering techniques in cardiac repair and disease modelling. Curr Pharm Des 20: 2048–56. doi: 10.2174/13816128113199990442
|
| [7] |
Sapir Y, Polyak B, Cohen S (2014) Cardiac tissue engineering in magnetically actuated scaffolds. Nanotechnology 25: 014009. doi: 10.1088/0957-4484/25/1/014009
|
| [8] | Williams C, Budina E, Stoppel WL, et al. (2014) Cardiac Extracellular Matrix-Fibrin Hybrid Scaffolds with Tunable Properties for Cardiovascular Tissue Engineering. Acta Biomater 14: 84–95. |
| [9] |
Galvez-Monton C, Prat-Vidal C, Roura S, et al. (2013) Cardiac Tissue Engineering and the Bioartificial Heart. Rev Esp Cardiol 66: 391–9. doi: 10.1016/j.recesp.2012.11.013
|
| [10] |
Roger VL, Go AS, Lloyd-Jones DM, et al. (2012) Heart disease and stroke statistics--2012 update: a report from the American Heart Association. Circulation 125: e2–e220. doi: 10.1161/CIR.0b013e31823ac046
|
| [11] | Tao ZW, Mohamed M, Hogan M, et al. (2014) Optimizing a spontaneously contracting heart tissue patch with rat neonatal cardiac cells on fibrin gel. J Tissue Eng Regen Med. doi: 10.1002/term.1895. |
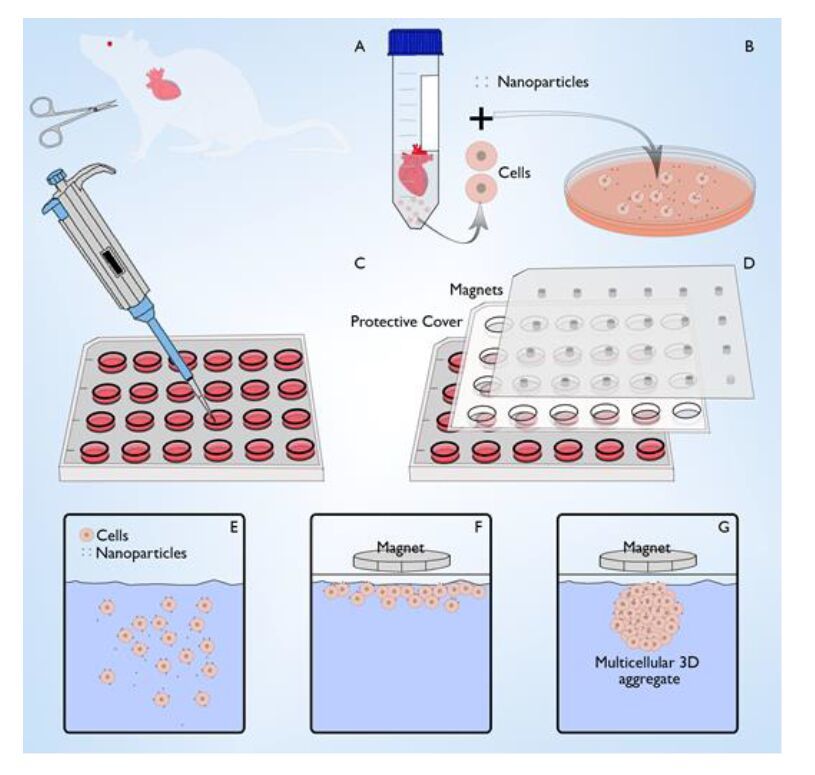

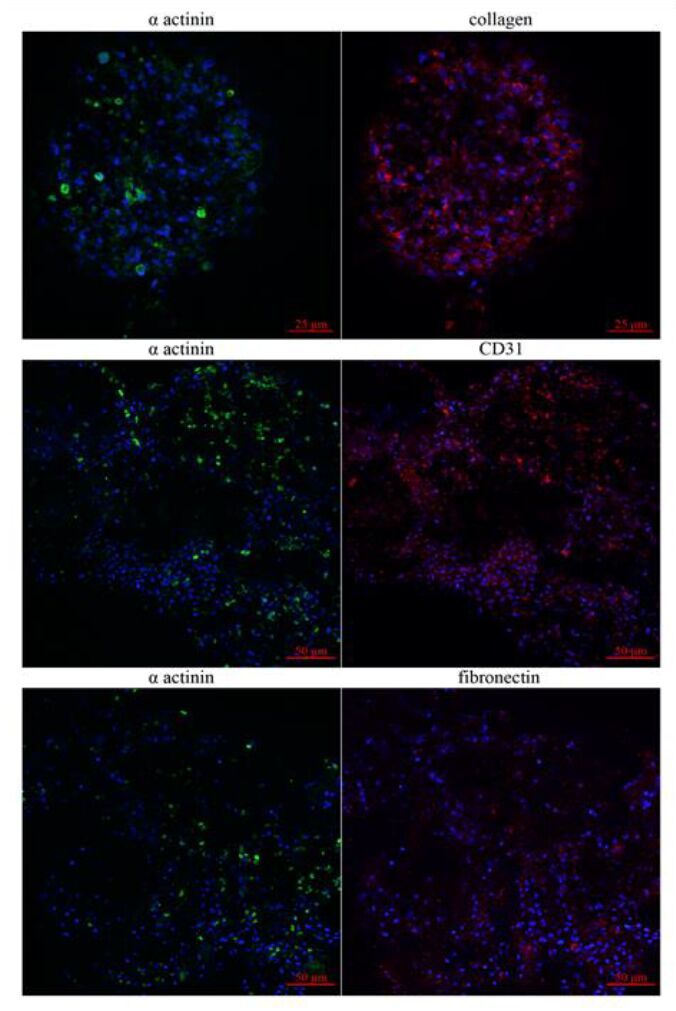
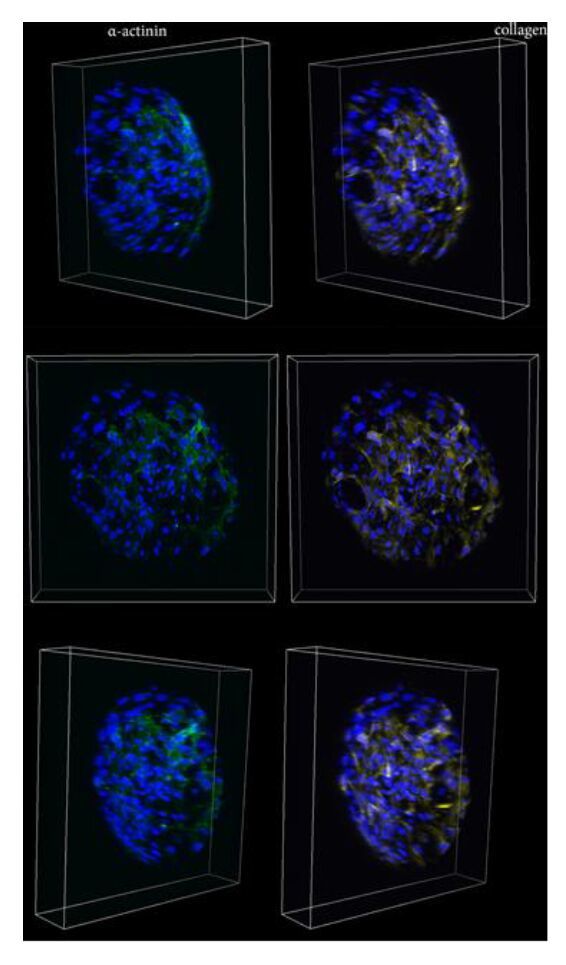
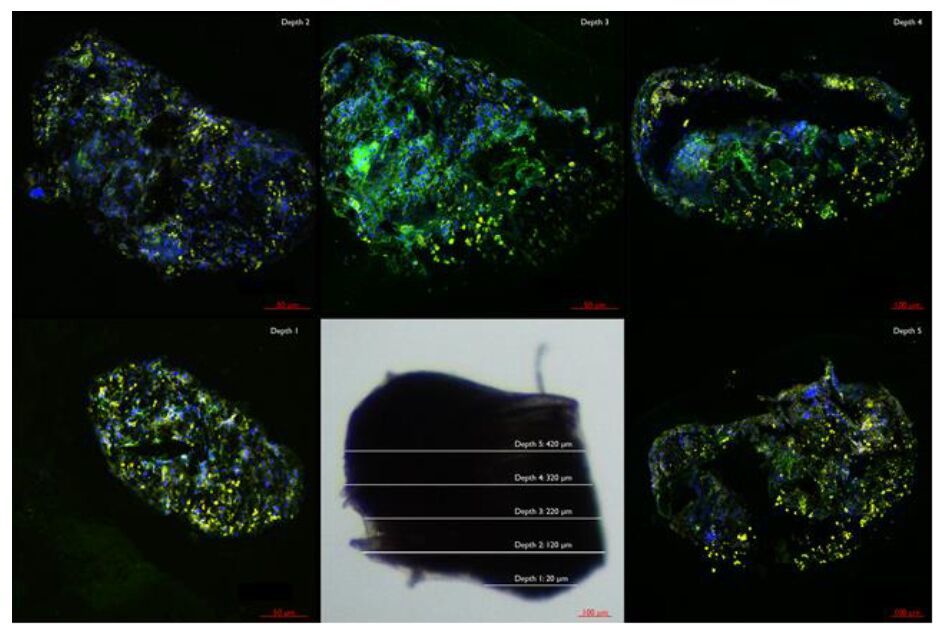
Figures(6)

Matthew Hogan, Glauco Souza, Ravi Birla. Assembly of a functional 3D primary cardiac construct using magnetic levitation[J]. AIMS Bioengineering, 2016, 3(3): 277-288. doi: 10.3934/bioeng.2016.3.277







 DownLoad:
DownLoad: