Citation: Qing Song, Yu Mao, Mark Wilkins, Fernando Segato, Rolf Prade. Cellulase immobilization on superparamagnetic nanoparticles for reuse in cellulosic biomass conversion[J]. AIMS Bioengineering, 2016, 3(3): 264-276. doi: 10.3934/bioeng.2016.3.264

| [1] |
Menon V, Rao M (2012) Trends in bioconversion of lignocellulose: Biofuels, platform chemicals &biorefinery concept. Prog Energy Combust Sci 38: 522–550. doi: 10.1016/j.pecs.2012.02.002

|
| [2] |
Wang H, Gurau G, Rogers RD (2012) Ionic liquid processing of cellulose. Chem Soc Rev 41: 1519–1537. doi: 10.1039/c2cs15311d
|
| [3] |
Bhat M, Bhat S (1997) Cellulose degrading enzymes and their potential industrial applications. Biotechnol Adv 15: 583–620. doi: 10.1016/S0734-9750(97)00006-2
|
| [4] |
Lynd LR, Weimer PJ, van Zyl WH, et al. (2002) Microbial cellulose utilization: fundamentals and biotechnology. Microbiol Mol Biol Rev 66: 506–577. doi: 10.1128/MMBR.66.3.506-577.2002
|
| [5] |
Lee SM, Jin LH, Kim JH, et al. (2010) β-glucosidase coating on polymer nanofibers for improved cellulosic ethanol production. Bioprocess Biosyst Eng 33: 141–147. doi: 10.1007/s00449-009-0386-x
|
| [6] |
Sheldon RA (2007) Enzyme immobilization: the quest for optimum performance. Adv Synth Catal 349: 1289–1307. doi: 10.1002/adsc.200700082
|
| [7] |
Mateo C, Palomo JM, Fernandez-Lorente G, et al. (2007) Improvement of enzyme activity, stability and selectivity via immobilization techniques. Enzyme Microb Technol 40: 1451–1463. doi: 10.1016/j.enzmictec.2007.01.018
|
| [8] |
Mohamad NR, Marzuki NHC, Buang NA, et al. (2015) An overview of technologies for immobilization of enzymes and surface analysis techniques for immobilized enzymes. Biotechnol Biotechnol Equip 29: 205–220. doi: 10.1080/13102818.2015.1008192
|
| [9] |
Chim-Anage P, Kashiwagi Y, Magae Y, et al. (1986) Properties of cellulase immobilized on agarose gel with spacer. Biotechnol Bioeng 28: 1876–1878. doi: 10.1002/bit.260281215
|
| [10] |
Isgrove F, Williams R, Niven G, et al. (2001) Enzyme immobilization on nylon-optimization and the steps used to prevent enzyme leakage from the support. Enzyme Microb Technol 28: 225–232. doi: 10.1016/S0141-0229(00)00312-4
|
| [11] |
Li C, Yoshimoto M, Fukunaga K, et al. (2007) Characterization and immobilization of liposome-bound cellulase for hydrolysis of insoluble cellulose. Bioresour Technol 98: 1366–1372. doi: 10.1016/j.biortech.2006.05.028
|
| [12] |
Georgelin T, Maurice V, Malezieux B, et al. (2010) Design of multifunctionalized γ-Fe2O3@SiO2 core–shell nanoparticles for enzymes immobilization. J Nanopart Res 12: 675–680. doi: 10.1007/s11051-009-9757-0
|
| [13] |
Wang P, Hu X, Cook S, et al. (2009) Influence of silica-derived nano-supporters on cellobiase after immobilization. Appl Biochem Biotechnol 158: 88–96. doi: 10.1007/s12010-008-8321-1
|
| [14] |
Verma ML, Chaudhary R, Tsuzuki T, et al. (2013) Immobilization of β-glucosidase on a magnetic nanoparticle improves thermostability: Application in cellobiose hydrolysis. Bioresour Technol 135: 2–6. doi: 10.1016/j.biortech.2013.01.047
|
| [15] |
Wood TM, Bhat KM (1988) Methods for measuring cellulase activities. Methods Enzymol 160: 87–112. doi: 10.1016/0076-6879(88)60109-1
|
| [16] | Saha BC, Bothast RJ (1996) Production, purification, and characterization of a highly glucose-tolerant novel beta-glucosidase from Candida peltata. Appl Environ Microbiol 62: 3165–3170. |
| [17] |
Segato F, Damásio AR, Gonçalves TA, et al. (2012) High-yield secretion of multiple client proteins in Aspergillus. Enzyme Microb Technol 51: 100–106. doi: 10.1016/j.enzmictec.2012.04.008
|
| [18] |
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72: 248–254. doi: 10.1016/0003-2697(76)90527-3
|
| [19] |
Shapiro AL, Viñuela E, Maizel JV Jr (1967) Molecular weight estimation of polypeptide chains by electrophoresis in SDS-polyacrylamide gels. Biochem Biophys Res Commun 28: 815–820. doi: 10.1016/0006-291X(67)90391-9
|
| [20] |
Caruntu D, Caruntu G, Chen Y, et al. (2004) Synthesis of variable-sized nanocrystals of Fe3O4 with high surface reactivity. Chem Mater 16: 5527–5534. doi: 10.1021/cm0487977
|
| [21] |
Lu Y, Yin Y, Mayers BT, et al. (2002) Modifying the surface properties of superparamagnetic iron oxide nanoparticles through a sol-gel approach. Nano Lett 2: 183–186. doi: 10.1021/nl015681q
|
| [22] | Caruntu D, Caruntu G, O'Connor CJ (2007) Magnetic properties of variable-sized Fe3O4 nanoparticles synthesized from non-aqueous homogeneous solutions of polyols. J Phys D Appl Phys 40: 5801–5809. |
| [23] | Wahajuddin, Arora S (2012) Superparamagnetic iron oxide nanoparticles: magnetic nanoplatforms as drug carriers. Int J Nanomedicine 7: 3445–3471. |
| [24] | Kannan K, Jasra RV (2010) Improved catalytic hydrolysis of carboxy methyl cellulose using cellulase immobilized on functionalized meso cellular foam. J Porous Mat 18: 409–416. |
| [25] | Jung C (2000) Insight into protein structure and protein–ligand recognition by Fourier transform infrared spectroscopy. J Mol Recognit 13: 325–351. |
| [26] | Gotić M, Musić S (2007) Mössbauer, FT-IR and FE SEM investigation of iron oxides precipitated from FeSO4 solutions. J Mol Struct 834: 445–453. |
| [27] | Gai L, Li Z, Hou Y, et al. (2010) Preparation of core-shell Fe3O4/SiO2 microspheres as adsorbents for purification of DNA. J Phys D Appl Phys 43: 5625–5634. |
| [28] |
Zhang D, Hegab HE, Lvov Y, et al. (2016) Immobilization of cellulase on a silica gel substrate modified using a 3-APTES self-assembled monolayer. Springerplus 5: 48. doi: 10.1186/s40064-016-1682-y
|
| [29] | Zhou J, Zhang J, Gao W (2014) Enhanced and selective delivery of enzyme therapy to 9L-glioma tumor via magnetic targeting of PEG-modified, β-glucosidase-conjugated iron oxide nanoparticles. Int J Nanomedicine 9: 2905–2917. |
| [30] |
Abraham RE, Verma ML, Barrow CJ, et al. (2014) Suitability of magnetic nanoparticle immobilised cellulases in enhancing enzymatic saccharification of pretreated hemp biomass. Biotechnol Biofuels 7: 90–101. doi: 10.1186/1754-6834-7-90
|
| [31] | Ladeira SA, Cruz E, Delatorre AB, et al. (2015). Cellulase production by thermophilic Bacillus sp. SMIA-2 and its detergent compatibility. Electron J Biotechn 18: 110–115. |
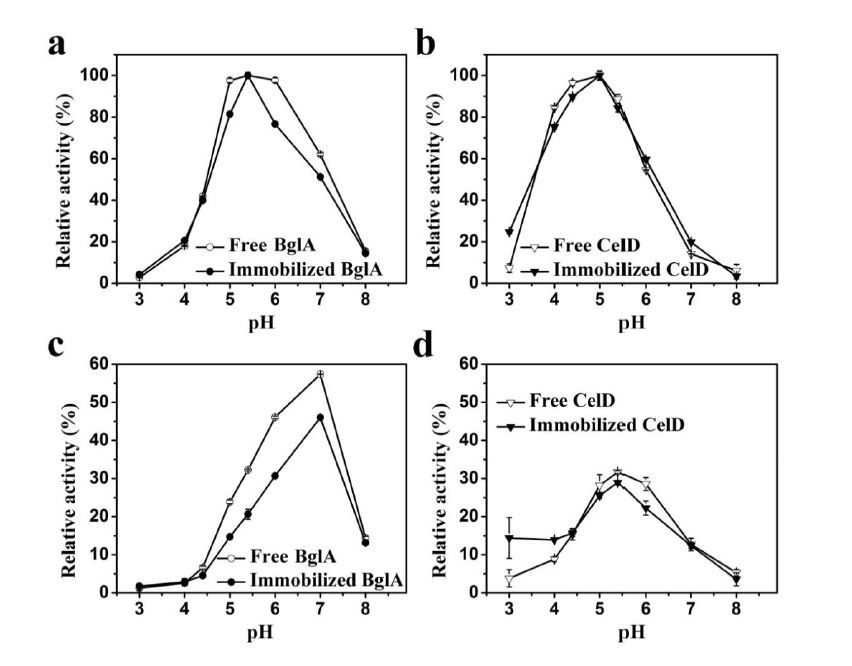
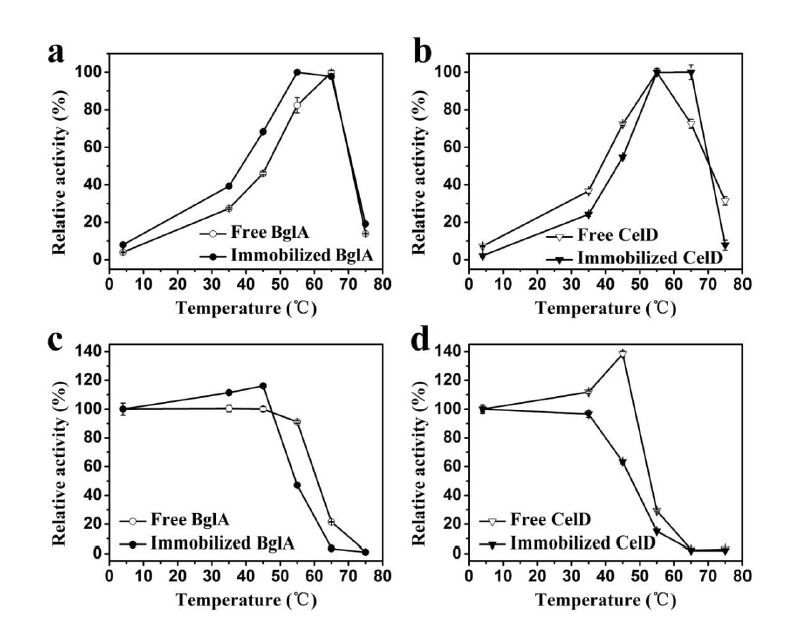
Figures(6)

Qing Song, Yu Mao, Mark Wilkins, Fernando Segato, Rolf Prade. Cellulase immobilization on superparamagnetic nanoparticles for reuse in cellulosic biomass conversion[J]. AIMS Bioengineering, 2016, 3(3): 264-276. doi: 10.3934/bioeng.2016.3.264







 DownLoad:
DownLoad: