Citation: Lucjan T. Orlowski, Monika Sywak. Wavering interactions between commodity futures prices and us dollar exchange rates[J]. Quantitative Finance and Economics, 2019, 3(2): 221-243. doi: 10.3934/QFE.2019.2.221

| [1] |
Akram QF (2009) Commodity prices, interest rates and the dollar. Energy Econ 31: 838–851. doi: 10.1016/j.eneco.2009.05.016

|
| [2] |
Allegret JP, Mignon V, Sallenave A (2015) Oil price shocks: Lessons from a model with trade and financial interdependencies. Econ Model 49: 232–247. doi: 10.1016/j.econmod.2015.04.009
|
| [3] |
Allegret JP, Couharde C, Mignon V, et al. (2017) Oil currencies in the face of oil shocks: What can be learned from time-varying specifications? Appl Econ 49: 1774–1793. doi: 10.1080/00036846.2016.1226490
|
| [4] |
Beckmann J, Czudaj R (2013) Oil prices and effective dollar exchange rates. Int Rev Econ Financ 27: 621–636. doi: 10.1016/j.iref.2012.12.002
|
| [5] | Bénassy-Quéré A, Mignon V, Penot A (2005) China and the relationship between the oil price and the dollar. CEPII Working Paper No. 2005–16. |
| [6] | Blanchard OJ, Gali J (2007) The macroeconomic effects of oil shocks: Why are the 2000s so different from the 1970s?. NBER Working Paper No. 13368. |
| [7] | Breitenfellner A, Crespo-Cuaresma J (2008) Crude oil prices and the USD/EUR exchange rate. Monetary Policy Econ Q 4. |
| [8] | Chen YC, Rogoff KS, Rossi B (2010) Can exchange rates forecast commodity prices? Q J Econ 125: 1145–1194. |
| [9] |
Chiang I-HE, Hughen WK, Sagi JS (2015) Estimating oil risk factors using information from equity and derivatives markets. J Financ 70: 769–804. doi: 10.1111/jofi.12222
|
| [10] |
Coudert V, Mignon V (2016) Reassessing the empirical relationship between the oil price and the dollar. Energy Policy 95: 147–157. doi: 10.1016/j.enpol.2016.05.002
|
| [11] |
Ding L, Vo M (2012) Exchange rates and oil prices: A multivariate volatility analysis. Q Rev Econ Financ 52: 15–37. doi: 10.1016/j.qref.2012.01.003
|
| [12] | Fratzscher M, Schneider D, van Robays I (2014) Oil prices, exchange rates and asset prices. European Central Bank Working Paper No. 1689. |
| [13] |
Joëts M, Mignon V, Razafindrabe T (2017) Does the volatility of commodity prices reflect macroeconomic uncertainty? Energy Econ 68: 313–326. doi: 10.1016/j.eneco.2017.09.017
|
| [14] |
Lizardo RA, Mollick AV (2010) Oil price fluctuations and US dollar exchange rates. Energy Econ 32: 399–408. doi: 10.1016/j.eneco.2009.10.005
|
| [15] |
Orlowski LT (2012) Financial crisis and extreme market risks: Evidence from Europe. Rev Financ Econ 21: 120–130. doi: 10.1016/j.rfe.2012.06.006
|
| [16] |
Orlowski LT (2015) Monetary expansion and bank credit: A lack of spark. J Policy Model 37: 510–520. doi: 10.1016/j.jpolmod.2015.03.013
|
| [17] | Orlowski LT (2017) Volatility of commodity futures prices and market-implied inflation expectations. J Int Financ Mark Inst Money 51: 133–141. |
| [18] |
Reboredo JC (2012) Modeling oil price and exchange rate co-movements. J Policy Model 34: 419–440. doi: 10.1016/j.jpolmod.2011.10.005
|
| [19] |
Sadorsky P (2000) The empirical relationship between energy futures prices and exchange rates. Energy Econ 22: 253–266. doi: 10.1016/S0140-9883(99)00027-4
|
| [20] |
Sari R, Hammoudeh S, Soytas U (2010) Dynamics of oil price, precious metal prices and exchange rate. Energy Econ 32: 351–362. doi: 10.1016/j.eneco.2009.08.010
|
| [21] |
Wu CC, Chung H, Chang YH (2012) The economic value of co-movement between oil price and exchange rate using copula-based GARCH models. Energy Econ 34: 270–282. doi: 10.1016/j.eneco.2011.07.007
|
| [22] | Zhang YJ (2013) The links between the price of oil and the value of US Dollar. Int J Energy Econ Policy 3: 341–351. |
| [23] |
Zhang YJ, Fan Y, Tsai HT, et al. (2008) Spillover effect of US dollar exchange rate on oil prices. J Policy Model 30: 973–991. doi: 10.1016/j.jpolmod.2008.02.002
|
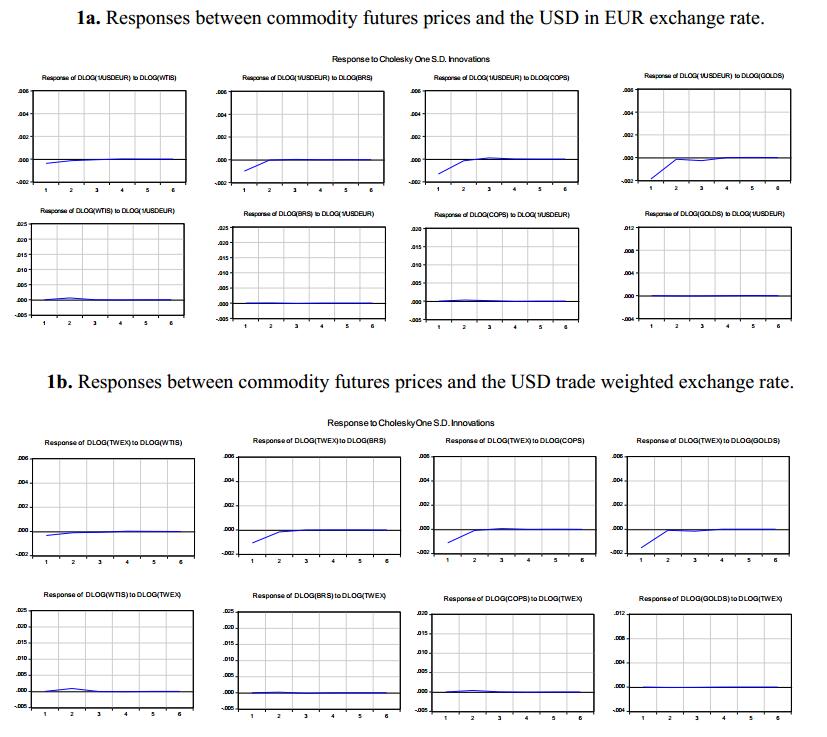
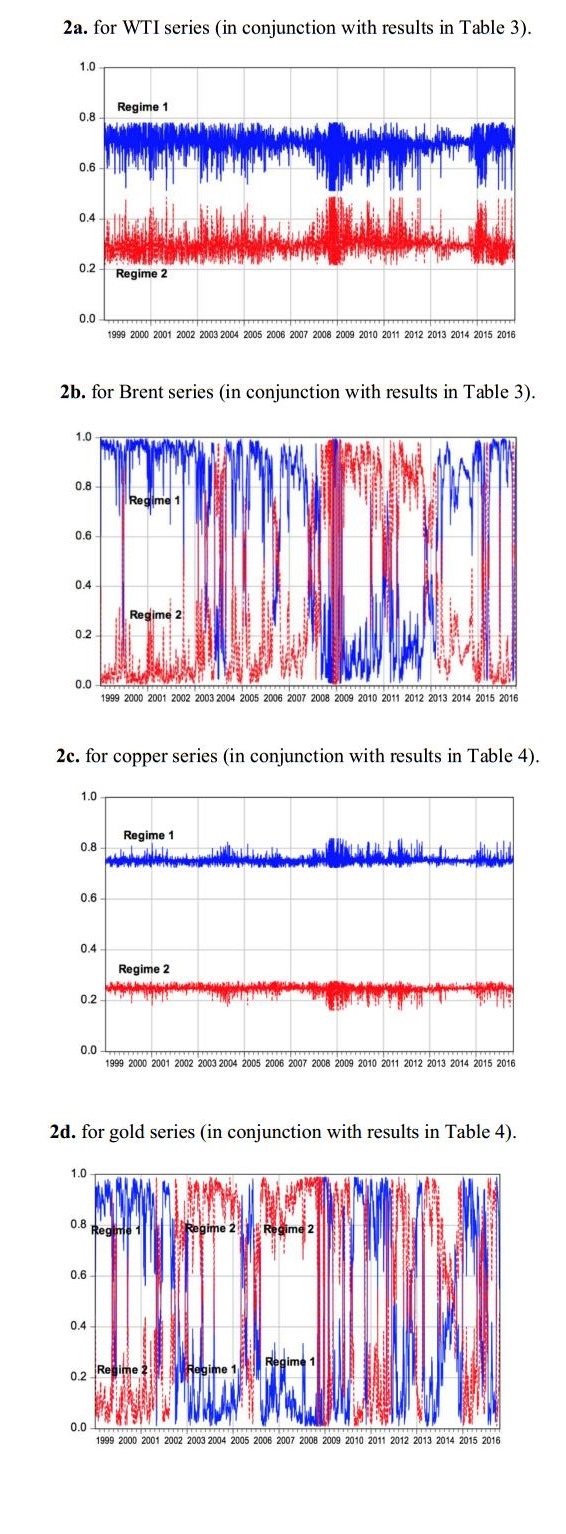

Figures(3) / Tables(8)

Lucjan T. Orlowski, Monika Sywak. Wavering interactions between commodity futures prices and us dollar exchange rates[J]. Quantitative Finance and Economics, 2019, 3(2): 221-243. doi: 10.3934/QFE.2019.2.221







 DownLoad:
DownLoad: