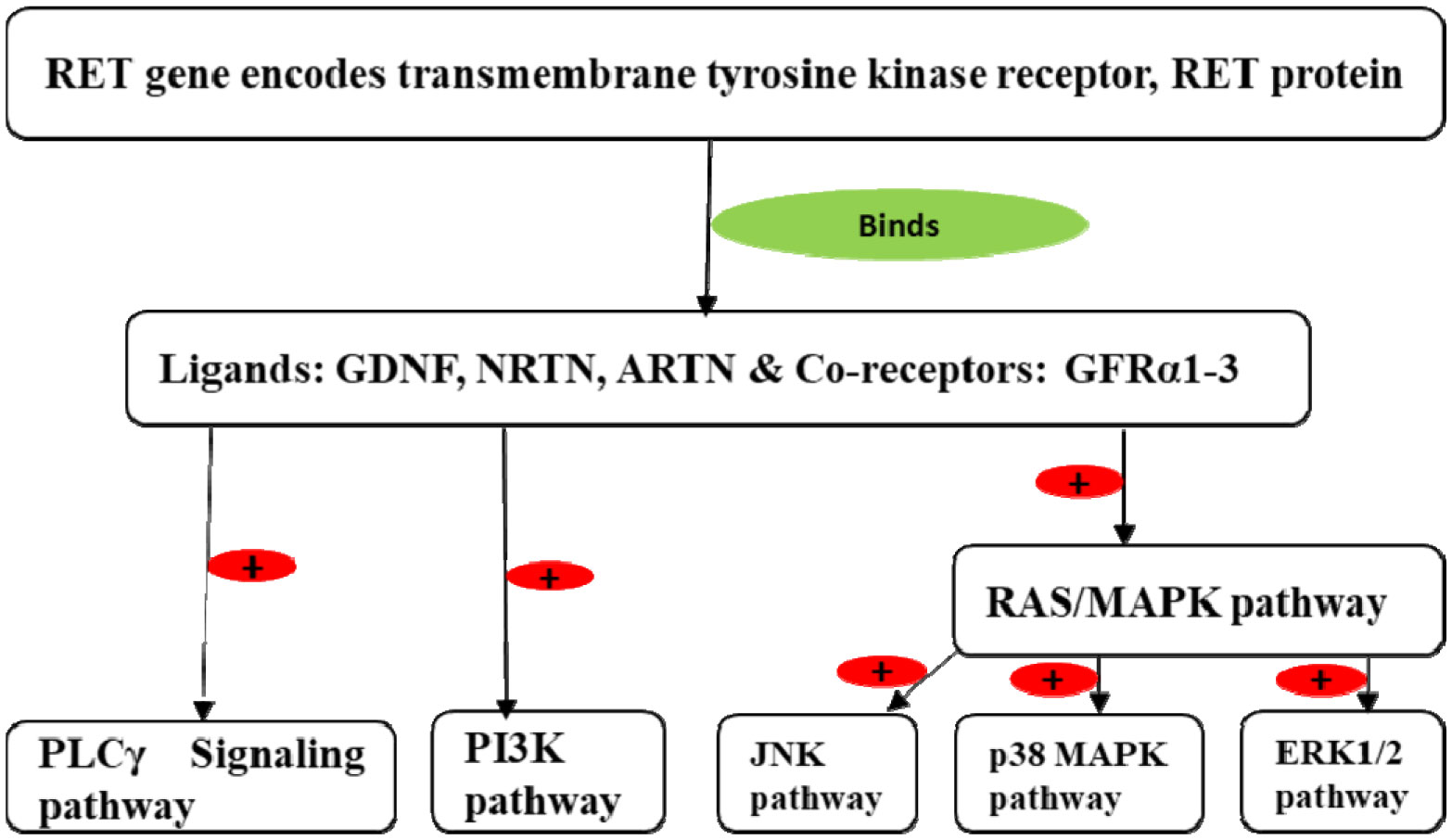
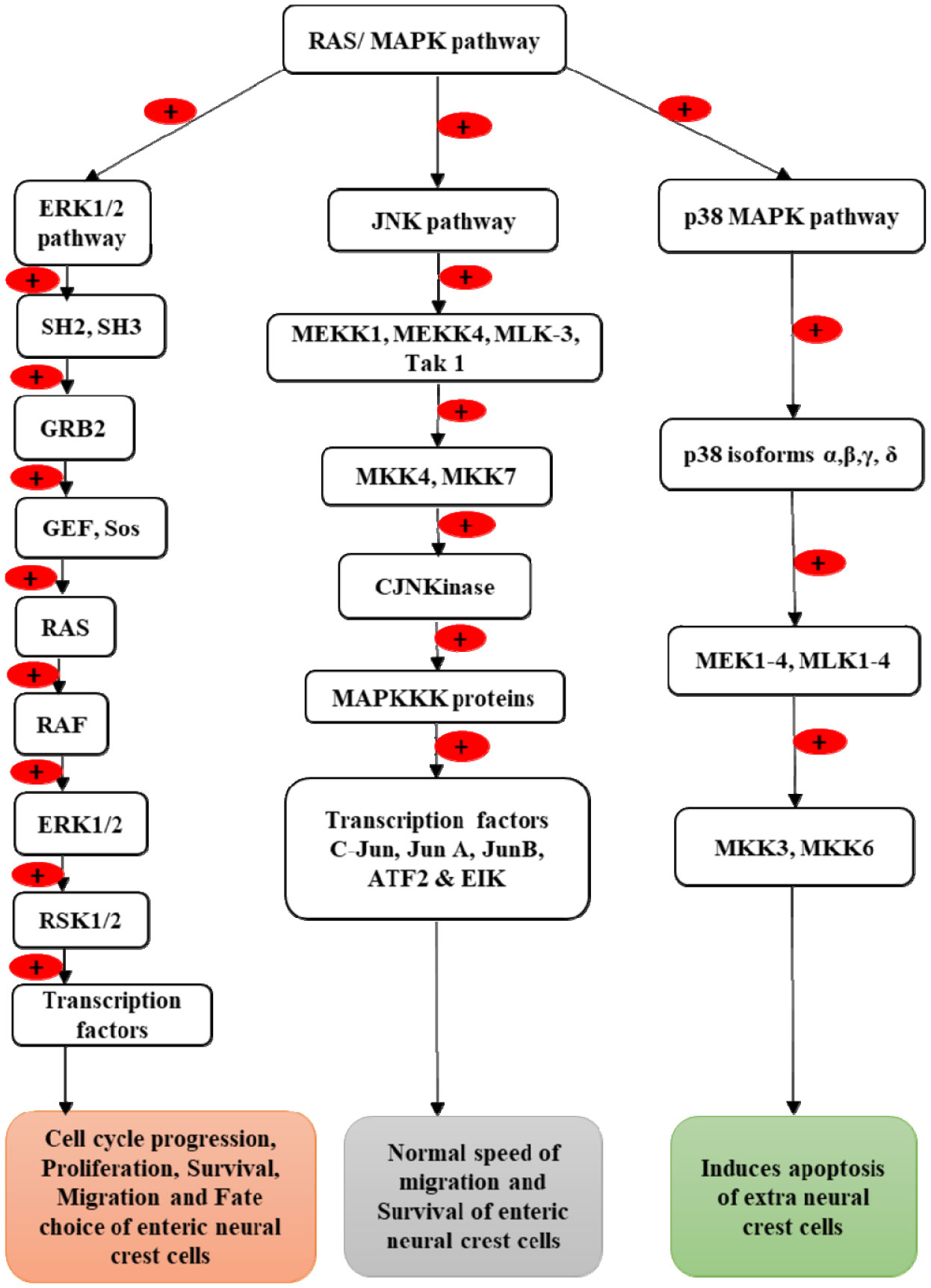
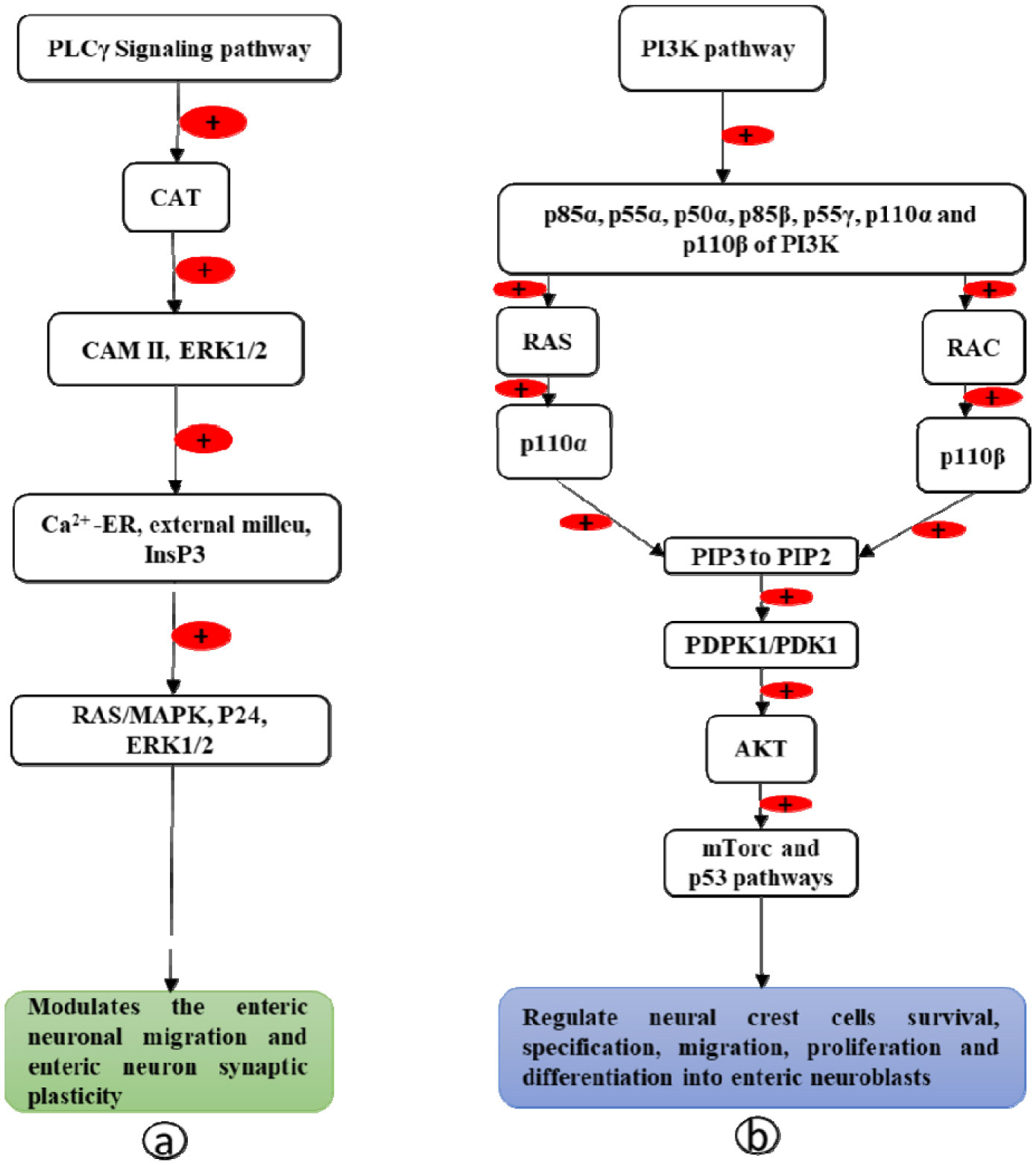
Enteric neurons and ganglia are derived from vagal and sacral neural crest cells, which undergo migration from the neural tube to the gut wall. In the gut wall, they first undergo rostrocaudal migration followed by migration from the superficial to deep layers. After migration, they proliferate and differentiate into the enteric plexus. Expression of the Rearranged During Transfection (RET) gene and its protein RET plays a crucial role in the formation of enteric neurons. This review describes the molecular mechanism by which the RET gene and the RET protein influence the development of enteric neurons. Vagal neural crest cells give rise to enteric neurons and glia of the foregut and midgut while sacral neural crest cells give rise to neurons of the hindgut. Interaction of RET protein with its ligands (glial cell derived neurotrophic factor (GDNF), neurturin (NRTN), and artemin (ARTN)) and its co-receptors (GDNF receptor alpha proteins (GFRα1-4)) activates the Phosphoinositide-3-kinase-protein kinase B (PI3K-PKB/AKT), RAS mitogen-activated protein kinase (RAS/MAPK) and phospholipase Cγ (PLCγ) signaling pathways, which control the survival, migration, proliferation, differentiation, and maturation of the vagal and sacral neural crest cells into enteric neurons. Abnormalities of the RET gene result in Hirschsprung's disease.
Citation: Chacchu Bhattarai, Phanindra Prasad Poudel, Arnab Ghosh, Sneha Guruprasad Kalthur. The RET gene encodes RET protein, which triggers intracellular signaling pathways for enteric neurogenesis, and RET mutation results in Hirschsprung's disease[J]. AIMS Neuroscience, 2022, 9(1): 128-149. doi: 10.3934/Neuroscience.2022008
Enteric neurons and ganglia are derived from vagal and sacral neural crest cells, which undergo migration from the neural tube to the gut wall. In the gut wall, they first undergo rostrocaudal migration followed by migration from the superficial to deep layers. After migration, they proliferate and differentiate into the enteric plexus. Expression of the Rearranged During Transfection (RET) gene and its protein RET plays a crucial role in the formation of enteric neurons. This review describes the molecular mechanism by which the RET gene and the RET protein influence the development of enteric neurons. Vagal neural crest cells give rise to enteric neurons and glia of the foregut and midgut while sacral neural crest cells give rise to neurons of the hindgut. Interaction of RET protein with its ligands (glial cell derived neurotrophic factor (GDNF), neurturin (NRTN), and artemin (ARTN)) and its co-receptors (GDNF receptor alpha proteins (GFRα1-4)) activates the Phosphoinositide-3-kinase-protein kinase B (PI3K-PKB/AKT), RAS mitogen-activated protein kinase (RAS/MAPK) and phospholipase Cγ (PLCγ) signaling pathways, which control the survival, migration, proliferation, differentiation, and maturation of the vagal and sacral neural crest cells into enteric neurons. Abnormalities of the RET gene result in Hirschsprung's disease.
Activating protein 1
Artemin
Activating transcription factor 2
Big MAP kinase
Calcium calmodulin-dependent kinase II
Cool-associated tyrosine-phosphorylated
C-Jun N-terminal kinase
Cadherin-like domain
E-twenty-six (ETS)-like transcription factor 1
E-twenty-six (ETS)-like transcription factor 7
Extracellular signal-regulated kinase
E 2 transcription factor
Glial cell-derived neurotrophic factor
Guanosine diphosphate
Guanine nucleotide exchange factor
Glial cell derived neurotrophic factor receptor alpha proteins 1-4
Glycosylphosphatidylinositol
Growth factor bound receptor protein 2
Guanosine triphosphate
Inositol 1,4,5-triphosphate receptor
Inositol 1,4,5-triphosphate
Jun N-terminal kinase
Mitogen activated protein kinase
Mitogen activated protein kinase kinases
Mitogen activated protein kinase kinase kinases
Myocyte enhancer transcription factor 2
Mitogen activated protein kinase-extracellular signal related kinase
Mitogen activated protein kinase kinase-extracellular signal related kinase kinase 1-4
Mitogen activated protein kinase kinase 3
Mitogen activated protein kinase kinase 4
Mitogen activated protein kinase kinase 6
Mitogen activated protein kinase kinase 7
Mixed lineage protein kinase 3
Mammalian target of rapamycin
Myelocytomatosis transcription factor
Neurturin
3-phosphoinositide-dependent protein kinase 1
Phosphatidylinositol (4,5)-bisphosphate
Phosphatidylinositol (3,4,5)-trisphosphate
Phosphoinositide-3-kinase
Protein kinase B
Phospholipase Cγ
Persephin
Phosphotyrosine-binding domain
p38 mitogen-activated protein kinase
Ras-related C3 botulinum toxin substrate
Rapidly accelerated fibrosarcoma
Rat Sarcoma Virus
Ras mitogen-activated protein kinase
Rearranged during transfection
Ribosomal S6 kinases
Serum- and glucocorticoid-inducible kinase
Src homolog 2
Src homolog 3
Son of sevenless
Specificity protein transcription factor 1
Transforming growth factor-β-activated kinase 1
Transforming growth factor-β
Tyrosine

| [1] |
Durbec PL, Larsson-Blomberg LB, Schuchardt A, et al. (1996) Common origin and developmental dependence on c-ret of subsets of enteric and sympathetic neuroblasts. Development 122: 349-358. https://doi.org/10.1242/dev.122.1.349 
|
| [2] |
Yntema CL, Hammond WS (1954) The origin of intrinsic ganglia of trunk viscera from vagal neural crest in the chick embryo. J Comp Neurol 101: 515-541. https://doi.org/10.1002/cne.901010212
|
| [3] |
Heanue TA, Pachnis V (2007) Enteric nervous system development and Hirschsprung's disease: Advances in genetic and stem cell studies. Nat Rev Neurosci 8: 466-479. https://doi.org/10.1038/nrn2137
|
| [4] |
Heuckeroth RO, Schäfer KH (2016) Gene-environment interactions and the enteric nervous system: Neural plasticity and Hirschsprung disease prevention. Dev Biol 417: 188-197. https://doi.org/10.1016/j.ydbio.2016.03.017
|
| [5] |
Tomuschat C, Puri P (2015) RET gene is a major risk factor for Hirschsprung's disease: a meta-analysis. Pediatr Surg Int 31: 701-710. https://doi.org/10.1007/s00383-015-3731-y
|
| [6] |
Taraviras S, Marcos-Gutierrez CV, Durbec P, et al. (1999) Signalling by the RET receptor tyrosine kinase and its role in the development of the mammalian enteric nervous system. Development 126: 2785-2797. https://doi.org/10.1242/dev.126.12.2785
|
| [7] | Le Douarin NM, Teillet MA (1973) The migration of neural crest cells to the wall of the digestive tract in avian embryo. J Embryol Exp Morphol 30: 31-48. https://doi.org/10.1242/dev.30.1.31 |
| [8] |
Kapur RP, Yost C, Palmiter RD (1992) A transgenic model for studying development of the enteric nervous system in normal and aganglionic mice. Development 116: 167-175. https://doi.org/10.1242/dev.116.1.167
|
| [9] |
Young HM, Hearn CJ, Ciampoli D, et al. (1998) A single rostrocaudal colonization of the rodent intestine by enteric neuron precursors is revealed by the expression of Phox2b, Ret, and p75 and by explants grown under the kidney capsule or in organ culture. Dev Biol 202: 67-84. https://doi.org/10.1006/dbio.1998.8987
|
| [10] |
Barlow AJ, Wallace AS, Thapar N, et al. (2008) Critical numbers of neural crest cells are required in the pathways from the neural tube to the foregut to ensure complete enteric nervous system formation. Development 135: 1681-1691. https://doi.org/10.1242/dev.017418
|
| [11] |
Barlow A, De Graaff E, Pachnis V (2003) Enteric Nervous System Progenitors Are Coordinately Controlled by the G Protein-Coupled Receptor EDNRB and the Receptor Tyrosine Kinase RET. Neuron 40: 905-916. https://doi.org/10.1016/S0896-6273(03)00730-X
|
| [12] |
Simpson MJ, Zhang DC, Mariani M, et al. (2007) Cell proliferation drives neural crest cell invasion of the intestine. Dev Biol 302: 553-568. https://doi.org/10.1016/j.ydbio.2006.10.017
|
| [13] |
Hao MM, Anderson RB, Kobayashi K, et al. (2009) The migratory behavior of immature enteric neurons. Dev Neurobiol 69: 22-35. https://doi.org/10.1002/dneu.20683
|
| [14] |
Hao MM, Young HM (2009) Development of enteric neuron diversity. J Cell Mol Med 13: 1193-1210. https://doi.org/10.1111/j.1582-4934.2009.00813.x
|
| [15] |
Jiang Y, Liu M tsai, Gershon MD (2003) Netrins and DCC in the guidance of migrating neural crest-derived cells in the developing bowel and pancreas. Dev Biol 258: 364-384. https://doi.org/10.1016/S0012-1606(03)00136-2
|
| [16] |
Burns AJ, Le Douarin NM (1998) The sacral neural crest contributes neurons and glia to the post-umbilical gut: Spatiotemporal analysis of the development of the enteric nervous system. Development 125: 4335-4347. https://doi.org/10.1242/dev.125.21.4335
|
| [17] | Newgreen D (2002) Enteric Nervous System: Development and Developmental Disturbances-Part 1 Understanding paediatric cancer (MEN2B and Neuroblastoma) using hESC and avian model View project Stem cell transplant in avian embryos View project. Springer . https://doi.org/10.1007/s10024001-0142-y |
| [18] |
Parisi MA, Kapur RP (2000) Genetics of Hirschsprung disease. Curr Opin Pediatr 12: 610-617. https://doi.org/10.1097/00008480-200012000-00017
|
| [19] |
Delalande JM, Barlow AJ, Thomas AJ, et al. (2008) The receptor tyrosine kinase RET regulates hindgut colonization by sacral neural crest cells. Dev Biol 313: 279-292. https://doi.org/10.1016/j.ydbio.2007.10.028
|
| [20] |
Schuchardt A, D'Agati V, Larsson-Blomberg L, et al. (1994) Defects in the kidney and enteric nervous system of mice lacking the tyrosine kinase receptor Ret. Nature 367: 380-383. https://doi.org/10.1038/367380a0
|
| [21] |
Marcos-Gutiérrez CV, Wilson SW, Holder N, et al. (1997) The zebrafish homologue of the ret receptor and its pattern of expression during embryogenesis. Oncogene 14: 879-889. https://doi.org/10.1038/sj.onc.1201048
|
| [22] |
Ceccherini I, Bocciardi R, Luo Y, et al. (1993) Exon structure and flanking intronic sequences of the human RET proto-oncogene. Biochem Bioph Res Co 196: 1288-1295. https://doi.org/10.1006/bbrc.1993.2392
|
| [23] |
Pasini B, Borrello MG, Greco A, et al. (1995) Loss of function effect of RET mutations causing Hirschsprung disease. Nat Genet 10: 35-40. https://doi.org/10.1038/ng0595-35
|
| [24] |
Romeo G, Ronchetto P, Luo Y, et al. (1994) Point mutations affecting the tyrosine kinase domain of the RET proto-oncogene in Hirschsprung's disease. Nature 367: 377-378. https://doi.org/10.1038/367377a0
|
| [25] |
Knowles PP, Murray-Rust J, Kjær S, et al. (2006) Structure and chemical inhibition of the RET tyrosine kinase domain. J Biol Chem 281: 33577-33587. https://doi.org/10.1074/jbc.M605604200
|
| [26] |
Takahashi M, Ritz J, Cooper GM (1985) Activation of a novel human transforming gene, ret, by DNA rearrangement. Cell 42: 581-588. https://doi.org/10.1016/0092-8674(85)90115-1
|
| [27] | Myers SM, Eng C, Ponder BAJ, et al. (1995) Characterization of RET proto-oncogene 3′ splicing variants and polyadenylation sites: A novel C-terminus for RET. Oncogene 11: 2039-2045. |
| [28] |
Natarajan D, Grigoriou M, Marcos-Gutierrez CV, et al. (1999) Multipotential progenitors of the mammalian enteric nervous system capable of colonising aganglionic bowel in organ culture. Development 126: 157-168. https://doi.org/10.1242/dev.126.1.157
|
| [29] |
Zhu L, Lee HO, Jordan CRS, et al. (2004) Spatiotemporal regulation of endothelin receptor-B by SOX10 in neural crest-derived enteric neuron precursors. Nat Genet 36: 732-737. https://doi.org/10.1038/ng1371
|
| [30] | RET - Proto-oncogene tyrosine-protein kinase receptor Ret precursor - Homo sapiens (Human) - RET gene & protein. Available from: https://www.uniprot.org/uniprot/P07949 |
| [31] |
Arighi E, Borrello MG, Sariola H (2005) RET tyrosine kinase signaling in development and cancer. Cytokine Growth F R 16: 441-467. https://doi.org/10.1016/j.cytogfr.2005.05.010
|
| [32] | Takahashi M, Buma Y, Hiai H (1989) Isolation of ret proto-oncogene cDNA with a an amino-terminal signal sequence. Oncogene 4: 805-806. |
| [33] | Takahashi M, Buma Y, Iwamoto T, et al. (1988) Cloning and expression of the ret proto-oncogene encoding a tyrosine kinase with two potential transmembrane domains. Oncogene 3: 571-578. |
| [34] | Kuma KI, Iwabe N, Miyata T (1993) Motifs of cadherin- and fibronectin type III-related sequences and evolution of the receptor-type-protein tyrosine kinases: Sequence similarity between proto-oncogene ret and cadherin family. Mol Biol Evol 10: 539-551. |
| [35] |
Anders J, Kjær S, Ibáñez CF (2001) Molecular Modeling of the Extracellular Domain of the RET Receptor Tyrosine Kinase Reveals Multiple Cadherin-like Domains and a Calcium-binding Site. J Biol Chem 276: 35808-35817. https://doi.org/10.1074/jbc.M104968200
|
| [36] |
Asai N, Iwashita T, Matsuyama M, et al. (1995) Mechanism of activation of the ret proto-oncogene by multiple endocrine neoplasia 2A mutations. Mol Cell Biol 15: 1613-1619. https://doi.org/10.1128/MCB.15.3.1613
|
| [37] |
Van Weering DHJ, Moen TC, Braakman I, et al. (1998) Expression of the receptor tyrosine kinase Ret on the plasma membrane is dependent on calcium. J Biol Chem 273: 12077-12081. https://doi.org/10.1074/jbc.273.20.12077
|
| [38] |
Liu X, Vega QC, Decker RA, et al. (1996) Oncogenic RET receptors display different autophosphorylation sites and substrate binding specificities. J Biol Chem 271: 5309-5312. https://doi.org/10.1074/jbc.271.10.5309
|
| [39] |
Kawamoto Y, Takeda K, Okuno Y, et al. (2004) Identification of RET Autophosphorylation Sites by Mass Spectrometry. J Biol Chem 279: 14213-14224. https://doi.org/10.1074/jbc.M312600200
|
| [40] |
Coulpier M, Anders J, Ibáñez CF (2002) Coordinated activation of autophosphorylation sites in the RET receptor tyrosine kinase: Importance of tyrosine 1062 for GDNF mediated neuronal differentiation and survival. J Biol Chem 277: 1991-1999. https://doi.org/10.1074/jbc.M107992200
|
| [41] |
Besset V, Scott RP, Ibáñez CF (2000) Signaling complexes and protein-protein interactions involved in the activation of the Ras and phosphatidylinositol 3-kinase pathways by the c-Ret receptor tyrosine kinase. J Biol Chem 275: 39159-39166. https://doi.org/10.1074/jbc.M006908200
|
| [42] |
Baloh RH, Enomoto H, Johnson EM, et al. (2000) The GDNF family ligands and receptors - Implications for neural development. Curr Opin Neurobiol 10: 103-110. https://doi.org/10.1016/S0959-4388(99)00048-3
|
| [43] |
Trupp M, Arenas E, Fainzilber M, et al. (1996) Functional receptor for GDNF encoded by the c-ret proto-oncogene. Nature 381: 785-789. https://doi.org/10.1038/381785a0
|
| [44] |
Kotzbauer PT, Lampe PA, Heuckeroth RO, et al. (1996) Neurturin, a relative of glial-cell-line-derived neurotrophic factor. Nature 384: 467-470. https://doi.org/10.1038/384467a0
|
| [45] |
Milbrandt J, De Sauvage FJ, Fahrner TJ, et al. (1998) Persephin, a novel neurotrophic factor related to GDNF and neurturin. Neuron 20: 245-253. https://doi.org/10.1016/S0896-6273(00)80453-5
|
| [46] |
Baloh RH, Tansey MG, Lampe PA, et al. (1998) Artemin, a novel member of the GDNF ligand family, supports peripheral and central neurons and signals through the GFRα3-RET receptor complex. Neuron 21: 1291-1302. https://doi.org/10.1016/S0896-6273(00)80649-2
|
| [47] |
Airaksinen MS, Titievsky A, Saarma M (1999) GDNF family neurotrophic factor signaling: Four masters, one servant. Mol Cell Neurosci 13: 313-325. https://doi.org/10.1006/mcne.1999.0754
|
| [48] |
Jing S, Wen D, Yu Y, et al. (1996) GDNF-induced activation of the Ret protein tyrosine kinase is mediated by GDNFR-α, a novel receptor for GDNF. Cell 85: 1113-1124. https://doi.org/10.1016/S0092-8674(00)81311-2
|
| [49] |
Trupp M, Raynoschek C, Belluardo N, et al. (1998) Multiple GPI-anchored receptors control GDNF-dependent and independent activation of the c-Ret receptor tyrosine kinase. Mol Cell Neurosci 11: 47-63. https://doi.org/10.1006/mcne.1998.0667
|
| [50] |
Amoresano A, Incoronato M, Monti G, et al. (2005) Direct interactions among Ret, GDNF and GFRα1 molecules reveal new insights into the assembly of a functional three-protein complex. Cell Signal 17: 717-727. https://doi.org/10.1016/j.cellsig.2004.10.012
|
| [51] |
Kjær S, Ibáñez CF (2003) Intrinsic susceptibility to misfolding of a hot-spot for Hirschsprung disease mutations in the ectodomain of RET. Hum Mol Genet 12: 2133-2144. https://doi.org/10.1093/hmg/ddg227
|
| [52] |
Kjær S, Hanrahan S, Totty N, et al. (2010) Mammal-restricted elements predispose human RET to folding impairment by HSCR mutations. Nat Struct Mol Biol 17: 726-731. https://doi.org/10.1038/nsmb.1808
|
| [53] |
Schlessinger J, Ullrich A (1992) Growth factor signaling by receptor tyrosine kinases. Neuron 9: 383-391. https://doi.org/10.1016/0896-6273(92)90177-F
|
| [54] |
Hayashi H, Ichihara M, Iwashita T, et al. (2000) Characterization of intracellular signals via tyrosine 1062 in RET activated by glial cell line-derived neurotrophic factor. Oncogene 19: 4469-4475. https://doi.org/10.1038/sj.onc.1203799
|
| [55] |
Segouffin-Cariou C, Billaud M (2000) Transforming ability of MEN2A-RET requires activation of the phosphatidylinositol 3-kinase/AKT signaling pathway. J Biol Chem 275: 3568-3576. https://doi.org/10.1074/jbc.275.5.3568
|
| [56] | Katz M, Amit I, Yarden Y (2007) Regulation of MAPKs by growth factors and receptor tyrosine kinases. BBA-Mol Cell Res 1773: 1161-1176. https://doi.org/10.1016/j.bbamcr.2007.01.002 |
| [57] |
Kim EK, Choi EJ (2010) Pathological roles of MAPK signaling pathways in human diseases. BBA-Molecular Basis Dis 1802: 396-405. https://doi.org/10.1016/j.bbadis.2009.12.009
|
| [58] |
Arighi E, Alberti L, Torriti F, et al. (1997) Identification of Shc docking site on Ret tyrosine kinase. Oncogene 14: 773-782. https://doi.org/10.1038/sj.onc.1200896
|
| [59] |
Pandey A, Liu X, Dixon JE, et al. (1996) Direct association between the Ret receptor tyrosine kinase and the Src homology 2-containing adapter protein Grb7. J Biol Chem 271: 10607-10610. https://doi.org/10.1074/jbc.271.18.10607
|
| [60] |
Lorenzo MJ, Gish GD, Houghton C, et al. (1997) RET alternate splicing influences the interaction of activated RET with the SH2 and PTB domains of Shc, and the SH2 domain of Grb2. Oncogene 14: 763-771. https://doi.org/10.1038/sj.onc.1200894
|
| [61] |
Li N, Batzer A, Daly R, et al. (1993) Guanine-nucleotide-releasing factor hSos1 binds to Grb2 and links receptor tyrosine kinases to Ras signalling. Nature 363: 85-88. https://doi.org/10.1038/363085a0
|
| [62] |
Roux PP, Blenis J (2004) ERK and p38 MAPK-Activated Protein Kinases: a Family of Protein Kinases with Diverse Biological Functions. Microbiol Mol Biol R 68: 320-344. https://doi.org/10.1128/MMBR.68.2.320-344.2004
|
| [63] |
Murphy LO, Blenis J (2006) MAPK signal specificity: the right place at the right time. Trends Biochem Sci 31: 268-275. https://doi.org/10.1016/j.tibs.2006.03.009
|
| [64] | Plotnikov A, Zehorai E, Procaccia S, et al. (2011) The MAPK cascades: Signaling components, nuclear roles and mechanisms of nuclear translocation. BBA-Mol Cell Res 1813: 1619-1633. https://doi.org/10.1016/j.bbamcr.2010.12.012 |
| [65] |
Matallanas D, Birtwistle M, Romano D, et al. (2011) Raf family kinases: Old dogs have learned new tricks. Genes and Cancer 2: 232-260. https://doi.org/10.1177/1947601911407323
|
| [66] |
Kato Y, Chao TH, Hayashi M, et al. (2000) Role of BMK1 in regulation of growth factor-induced cellular responses. Immunol Res 21: 233-237. https://doi.org/10.1385/IR:21:2-3:233
|
| [67] |
Watson FL, Heerssen HM, Bhattacharyya A, et al. (2001) Neurotrophins use the Erk5 pathway to mediate a retrograde survival response. Nat Neurosci 4: 981-988. https://doi.org/10.1038/nn720
|
| [68] |
Cargnello M, Roux PP (2011) Activation and Function of the MAPKs and Their Substrates, the MAPK-Activated Protein Kinases. Microbiol Mol Biol R 75: 50-83. https://doi.org/10.1128/MMBR.00031-10
|
| [69] |
Wortzel I, Seger R (2011) The ERK cascade: Distinct functions within various subcellular organelles. Genes and Cancer 2: 195-209. https://doi.org/10.1177/1947601911407328
|
| [70] |
Davis RJ (2000) Signal transduction by the JNK group of MAP kinases. Cell 103: 239-252. https://doi.org/10.1016/S0092-8674(00)00116-1
|
| [71] |
Kyriakis JM, Avruch J (2001) Mammalian mitogen-activated protein kinase signal transduction pathways activated by stress and inflammation. Physiol Rev 81: 807-869. https://doi.org/10.1152/physrev.2001.81.2.807
|
| [72] |
Teramoto H, Coso OA, Miyata H, et al. (1996) Signaling from the small GTP-binding proteins Rac1 and Cdc42 to the c- Jun N-terminal kinase/stress-activated protein kinase pathway: A role for mixed lineage kinase 3/protein-tyrosine kinase 1, a novel member of the mixed lineage kinase family. J Biol Chem 271: 27225-27228. https://doi.org/10.1074/jbc.271.44.27225
|
| [73] |
Minden A, Lin A, Claret FX, et al. (1995) Selective activation of the JNK signaling cascadeand c-Jun transcriptional activity by the small GTPases Rac and Cdc42Hs. Cell 81: 1147-1157. https://doi.org/10.1016/S0092-8674(05)80019-4
|
| [74] |
Fanger GR, Johnson NL, Johnson GL (1997) MEK kinases are regulated by EGF and selectively interact with Rac/Cdc42. EMBO J 16: 4961-4972. https://doi.org/10.1093/emboj/16.16.4961
|
| [75] |
Vincentz JW, Barnes RM, Rodgers R, et al. (2008) An absence of Twist1 results in aberrant cardiac neural crest morphogenesis. Dev Biol 320: 131-139. https://doi.org/10.1016/j.ydbio.2008.04.037
|
| [76] |
Vincentz JW, Firulli BA, Lin A, et al. (2013) Twist1 Controls a Cell-Specification Switch Governing Cell Fate Decisions within the Cardiac Neural Crest. PLoS Genet 9. https://doi.org/10.1371/journal.pgen.1003405
|
| [77] | Yan C, Liang Y, Nylander KD, et al. (2002) TrkA as a life and death receptor: Receptor dose as a mediator of function. Cancer Res 62: 4867-4875. |
| [78] |
Rhee SG, Suh PG, Ryu SH, et al. (1989) Studies of inositol phospholipid - Specific phospholipase C. Science 244: 546-550. https://doi.org/10.1126/science.2541501
|
| [79] |
Rhee Sue Goo, Bae Yun Soo (1997) Regulation of phosphoinositide-specific phospholipase C isozymes. J Biol Chem 272: 15045-15048. https://doi.org/10.1074/jbc.272.24.15045
|
| [80] |
Borrello MG, Alberti L, Arighi E, et al. (1996) The full oncogenic activity of Ret/ptc2 depends on tyrosine 539, a docking site for phospholipase Cgamma. Mol Cell Biol 16: 2151-2163. https://doi.org/10.1128/MCB.16.5.2151
|
| [81] |
Arneson LN, Leibson PJ (2005) Signaling in natural immunity: Natural killer cells. NeuroImmune Biology . Elsevier 151-166. https://doi.org/10.1016/S1567-7443(05)80014-6
|
| [82] |
Rusciano MR, Salzano M, Monaco S, et al. (2010) The Ca2+-calmodulin-dependent kinase II is activated in papillary thyroid carcinoma (PTC) and mediates cell proliferation stimulated by RET/PTC. Endocr-Relat Cancer 17: 113-123. https://doi.org/10.1677/ERC-09-0214
|
| [83] |
Lundgren TK, Nakahata K, Fritz N, et al. (2012) RET PLCγ phosphotyrosine binding domain regulates CA 2+ signaling and neocortical neuronal migration. PLoS ONE 7: e31258. https://doi.org/10.1371/journal.pone.0031258
|
| [84] |
Berridge MJ, Bootman MD, Roderick HL (2003) Calcium signalling: Dynamics, homeostasis and remodelling. Nat Rev Mol Cell Bio 4: 517-529. https://doi.org/10.1038/nrm1155
|
| [85] |
Uhlén P, Fritz N (2010) Biochemistry of calcium oscillations. Biochem Bioph Res Co 396: 28-32. https://doi.org/10.1016/j.bbrc.2010.02.117
|
| [86] |
Desfrere L, Karlsson M, Hiyoshi H, et al. (2009) Na,K-ATPase signal transduction triggers CREB activation and dendritic growth. P Natl Acad Sci USA 106: 2212-2217. https://doi.org/10.1073/pnas.0809253106
|
| [87] |
Fruman DA, Chiu H, Hopkins BD, et al. (2017) The PI3K Pathway in Human Disease. Cell 170: 605-635. https://doi.org/10.1016/j.cell.2017.07.029
|
| [88] |
Fritsch R, De Krijger I, Fritsch K, et al. (2013) XRAS and RHO families of GTPases directly regulate distinct phosphoinositide 3-kinase isoforms. Cell 153: 1050. https://doi.org/10.1016/j.cell.2013.04.031
|
| [89] |
Maehama T, Dixon JE (1998) The tumor suppressor, PTEN/MMAC1, dephosphorylates the lipid second messenger, phosphatidylinositol 3,4,5-trisphosphate. J Biol Chem 273: 13375-13378. https://doi.org/10.1074/jbc.273.22.13375
|
| [90] |
Sarbassov DD, Guertin DA, Ali SM, et al. (2005) Phosphorylation and regulation of Akt/PKB by the rictor-mTOR complex. Science 307: 1098-1101. https://doi.org/10.1126/science.1106148
|
| [91] |
Alessi DR, James SR, Downes CP, et al. (1997) Characterization of a 3-phosphoinositide-dependent protein kinase which phosphorylates and activates protein kinase Bα. Curr Biol 7: 261-269. https://doi.org/10.1016/S0960-9822(06)00122-9
|
| [92] |
Stokoe D, Stephens LR, Copeland T, et al. (1997) Dual role of phosphatidylinositol-3,4,5-trisphosphate in the activation of protein kinase B. Science 277: 567-570. https://doi.org/10.1126/science.277.5325.567
|
| [93] |
Focke PJ, Schiltz CA, Jones SE, et al. (2001) Enteric neuroblasts require the phosphatidylinositol 3-kinase pathway for GDNF-stimulated proliferation. J Neurobiol 47: 306-317. https://doi.org/10.1002/neu.1037
|
| [94] |
Manning BD, Toker A (2017) AKT/PKB Signaling: Navigating the Network. Cell 169: 381-405. https://doi.org/10.1016/j.cell.2017.04.001
|
| [95] |
Jhiang SM (2000) The RET proto-oncogene in human cancers. Oncogene 19: 5590-5597. https://doi.org/10.1038/sj.onc.1203857
|
| [96] |
Eng C (1999) RET proto-oncogene in the development of human cancer. J Clin Oncol 1: 380-393. https://doi.org/10.1200/JCO.1999.17.1.380
|
| [97] |
Mulligan LM, Eng C, Attlé T, et al. (1994) Diverse phenotypes associated with exon 10 mutations of the RET proto-oncogene. Hum Mol Genet 3: 2163-2168. https://doi.org/10.1093/hmg/3.12.2163
|
| [98] |
Edery P, Lyonnet S, Mulligan LM, et al. (1994) Mutations of the RET proto-oncogene in Hirschsprung's disease. Nature 367: 378-380. https://doi.org/10.1038/367378a0
|
| [99] |
Angrist M, Bolk S, Thiel B, et al. (1995) Mutation analysis of the RET receptor tyrosine kinase in hirschsprung disease. Hum Mol Genet 4: 821-830. https://doi.org/10.1093/hmg/4.5.821
|
| [100] |
Eng C, Mulligan LM (1997) Mutations of the RET proto-oncogene in the multiple endocrine neoplasia type 2 syndromes, related sporadic tumours, and Hirschsprung disease. Hum Mutat 9: 97-109. https://doi.org/10.1002/(SICI)1098-1004(1997)9:2<97::AID-HUMU1>3.0.CO;2-M
|
| [101] |
Jing S, Wen D, Yu Y, et al. (1996) GDNF-induced activation of the Ret protein tyrosine kinase is mediated by GDNFR-α, a novel receptor for GDNF. Cell 85: 1113-1124. https://doi.org/10.1016/S0092-8674(00)81311-2
|
| [102] |
Wu W, Lu L, Xu W, et al. (2019) Whole Exome Sequencing Identifies a Novel Pathogenic RET Variant in Hirschsprung Disease. Front Genet 9: 752. https://doi.org/10.3389/fgene.2018.00752
|
| [103] |
Yin L, Barone V, Seri M, et al. (1994) Heterogeneity and low detection rate of RET mutations in Hirschsprung disease. Eur J Hum Genet 2: 272-280. https://doi.org/10.1159/000472371
|
| [104] |
Iwashita T, Kurokawa K, Qiao S, et al. (2001) Functional analysis of RET with Hirschsprung mutations affecting its kinase domain. Gastroenterology 121: 24-33. https://doi.org/10.1053/gast.2001.25515
|
| [105] |
Fusco A, Grieco M, Santoro M, et al. (1988) A new oncogene in human thyroid papillary carcinomas and their lymph-nodal metastases. Nature 328: 170-172. https://doi.org/10.1038/328170a0
|
| [106] |
Van Heyningen V (1994) One gene - Four syndromes. Nature 367: 319-320. https://doi.org/10.1038/367319a0
|
| [107] |
Gath R, Goessling A, Keller KM, et al. (2001) Analysis of the RET, GDNF, EDN3, and EDNRB genes in patients with intestinal neuronal dysplasia and Hirschsprung disease. Gut 48: 671-675. https://doi.org/10.1136/gut.48.5.671
|
| [108] |
Julies MG, Moore SW, Kotze MJ, et al. (2001) Novel RET mutations in Hirschsprung's disease patients from the diverse South African population. Eur J Hum Genet 9: 419-423. https://doi.org/10.1038/sj.ejhg.5200650
|
| [109] |
Passarge E (1967) The Genetics of Hirschsprung's Disease. New Engl J Med 276: 138-143. https://doi.org/10.1056/NEJM196701192760303
|
| [110] |
Martucciello G, Bicocchi MP, Dodero P, et al. (1992) Total colonic aganglionosis associated with interstitial deletion of the long arm of chromosome 10. Pediatr Surg Int 7: 308-310. https://doi.org/10.1007/BF00183991
|
| [111] |
Lyonnet S, Bolino A, Pelet A, et al. (1993) A gene for Hirschsprung disease maps to the proximal long arm of chromosome 10. Nat Genet 4: 346-350. https://doi.org/10.1038/ng0893-346
|
| [112] |
Puliti A, Covone AE, Bicocchi MP, et al. (1993) Deleted and normal chromosome 10 homologs from a patient with Hirschsprung disease isolated in two cell hybrids through enrichment by immunomagnetic selection. Cytogenet Cell Genet 63: 102-106. https://doi.org/10.1159/000133510
|
| [113] |
Cosma MP, Cardone M, Carlomagno F, et al. (1998) Mutations in the Extracellular Domain Cause RET Loss of Function by a Dominant Negative Mechanism. Mol Cell Biol 18: 3321-3329. https://doi.org/10.1128/MCB.18.6.3321
|
| [114] |
Iwashita T, Murakarni H, Asai N, et al. (1996) Mechanism of Ret dysfunction by Hirschsprung mutations affecting its extracellular domain. Hum Mol Genet 5: 1577-1580. https://doi.org/10.1093/hmg/5.10.1577
|
| [115] |
Carlomagno F, De Vita G, Berlingieri MT, et al. (1996) Molecular heterogeneity of RET loss of function in Hirschsprung's disease. EMBO Journal 15: 2717-2725. https://doi.org/10.1002/j.1460-2075.1996.tb00632.x
|
| [116] |
Pelet A, Attie T, Goulet O, et al. (1994) De-novo mutations of the RET proto-oncogene in Hirschsprung's disease. The Lancet 344: 1769-1770. https://doi.org/10.1016/S0140-6736(94)92908-4
|
| [117] |
Attié T, Pelet A, Edery P, et al. (1995) Diversity of RET proto-oncogene mutations in familial and sporadic hirschsprung disease. Hum Mol Genet 4: 1381-1386. https://doi.org/10.1093/hmg/4.8.1381
|
| [118] | Chakravarti A (1996) Endothelin receptor-mediated signaling in Hirschsprung disease. Hum Mol Genet 5: 303-307. |
| [119] |
Seri M, Yin L, Barone V, et al. (1997) Frequency of Ret mutations in long- and short-segment Hirschsprung disease. Hum Mutat 9: 243-249. https://doi.org/10.1002/(SICI)1098-1004(1997)9:3<243::AID-HUMU5>3.0.CO;2-8
|
| [120] |
Geneste O, Bidaud C, De Vita G, et al. (1999) Two distinct mutations of the RET receptor causing Hirschsprung's disease impair the binding of signalling effectors to a multifunctional docking site. Hum Mol Genet 8: 1989-1999. https://doi.org/10.1093/hmg/8.11.1989
|
| [121] |
Treanor JJS, Goodman L, De Sauvage F, et al. (1996) Characterization of a multicomponent receptor for GDNF. Nature 382: 80-83. https://doi.org/10.1038/382080a0
|
| [122] |
Auricchio A, Griseri P, Carpentieri ML, et al. (1999) Double heterozygosity for a RET substitution interfering with splicing and an EDNRB missense mutation in Hirschsprung disease [3]. Am J Hum Genet 64: 1216-1221. https://doi.org/10.1086/302329
|
| [123] | Wang H, Li Q, Zhang Z, et al. (2019) Functional Studies on Novel RET Mutations and Their Implications for Genetic Counseling for Hirschsprung Disease. Front Genet 10. https://doi.org/10.3389/fgene.2019.00924 |
| [124] |
De Graaff E, Srinivas S, Kilkenny C, et al. (2001) Differential activities of the RET tyrosine kinase receptor isoforms during mammalian embryogenesis. Gene Dev 15: 2433-2444. https://doi.org/10.1101/gad.205001
|
| [125] |
Wong A, Bogni S, Kotka P, et al. (2005) Phosphotyrosine 1062 Is Critical for the In Vivo Activity of the Ret9 Receptor Tyrosine Kinase Isoform. Mol Cell Biol 25: 9661-9673. https://doi.org/10.1128/MCB.25.21.9661-9673.2005
|
| [126] |
Núñez-Torres R, Fernández RM, Acosta MJ, et al. (2011) Comprehensive analysis of RET common and rare variants in a series of Spanish Hirschsprung patients confirms a synergistic effect of both kinds of events. BMC Med Genet 12: 138. https://doi.org/10.1186/1471-2350-12-138
|
| [127] |
Torroglosa A, Alves MM, Fernández RM, et al. (2016) Epigenetics in ENS development and Hirschsprung disease. Dev Biol 417: 209-216. https://doi.org/10.1016/j.ydbio.2016.06.017
|
| [128] |
Carniti C, Belluco S, Riccardi E, et al. (2006) The RetC620R mutation affects renal and enteric development in a mouse model of Hirschsprung's disease. Am J Pathol 168: 1262-1275. https://doi.org/10.2353/ajpath.2006.050607
|
| [129] |
Jijiwa M, Fukuda T, Kawai K, et al. (2004) A Targeting Mutation of Tyrosine 1062 in Ret Causes a Marked Decrease of Enteric Neurons and Renal Hypoplasia. Mol Cell Biol 24: 8026-8036. https://doi.org/10.1128/MCB.24.18.8026-8036.2004
|
| [130] |
Shimotake T, Go S, Inoue K, et al. (2001) A homozygous missense mutation in the tyrosine kinase domain of the RET proto-oncogene in an infant with total intestinal aganglionosis. Am J Gastroenterol 96: 1286-1291. https://doi.org/10.1111/j.1572-0241.2001.03714.x
|
Figures(3) / Tables(11)

Chacchu Bhattarai, Phanindra Prasad Poudel, Arnab Ghosh, Sneha Guruprasad Kalthur. The RET gene encodes RET protein, which triggers intracellular signaling pathways for enteric neurogenesis, and RET mutation results in Hirschsprung's disease[J]. AIMS Neuroscience, 2022, 9(1): 128-149. doi: 10.3934/Neuroscience.2022008







 DownLoad:
DownLoad: