Citation: MD Main Uddin, Karen P Briski. Neuroestradiol regulation of ventromedial hypothalamic nucleus 5′-AMP-activated protein kinase activity and counterregulatory hormone secretion in hypoglycemic male versus female rats[J]. AIMS Neuroscience, 2021, 8(1): 133-147. doi: 10.3934/Neuroscience.2021006
5′-AMP-activated protein kinase
estrogen receptor
insulin
insulin-induced hypoglycemia
lateral ventricle
letrozole
mediobasal hypothalamus
ovariectomy
phosphoAMPK
ventromedial hypothalamic nucleus

| [1] |
Xue B, Kahn BB (2006) AMPK integrates nutrient and hormonal signals to regulate food intake and energy balance through effects in the hypothalamus and peripheral tissues. J Physiol 574: 73-83. doi: 10.1113/jphysiol.2006.113217

|
| [2] |
Pimentel GD, Ropelle ER, Rocha GZ, et al. (2013) The role of neuronal AMPK as a mediator of nutritional regulation of food intake and energy homeostasis. Metabolism 62: 171-178. doi: 10.1016/j.metabol.2012.07.001
|
| [3] |
López M (2018) Hypothalamic AMPK and energy balance. Eur J Clin Invest 48: e12996. doi: 10.1111/eci.12996
|
| [4] |
Ronnett GV, Ramamurthy S, Kleman AM, et al. (2009) AMPK in the brain: its roles in energy balance and neuroprotection. J Neurochem 109: 17-23. doi: 10.1111/j.1471-4159.2009.05916.x
|
| [5] | Huynh MKQ, Kinyua AW, Yang DJ, et al. (2016) Hypothalamic AMPK as a Regulator of Energy Homeostasis. Neural Plast 2016: 2754078. |
| [6] |
Han SM, Namkoong C, Jang PG, et al. (2005) Hypothalamic AMP-activated protein kinase mediates counter-regulatory responses to hypoglycaemia in rats. Diabetologia 48: 2170-2178. doi: 10.1007/s00125-005-1913-1
|
| [7] |
McCrimmon RJ, Shaw M, Fan X, et al. (2008) Key role for AMP-activated protein kinase in the ventromedial hypothalamus in regulating counterregulatory hormone responses to acute hypoglycemia. Diabetes 2008; 57: 444-450. doi: 10.2337/db07-0837
|
| [8] |
Ibrahim MMH, Bheemanapally K, Alhamami HN, et al. (2020) Effects of intracerebroventricular glycogen phosphorylase inhibitor CP-316, 819 infusion on hypothalamic glycogen content and metabolic neuron AMPK activity and neurotransmitter expression in the male rat. J Mol Neurosci 70: 647-658. doi: 10.1007/s12031-019-01471-0
|
| [9] |
Briski KP, Mandal SK, Bheemanapally K, et al. (2020) Effects of acute versus recurrent insulin-induced hypoglycemia on ventromedial hypothalamic nucleus metabolic-sensory neuron AMPK activity: Impact of alpha1-adrenergic receptor signaling. Brain Res Bull 157: 41-50. doi: 10.1016/j.brainresbull.2020.01.013
|
| [10] |
Nedungadi TP, Briski KP (2012) Site-specific effects of intracranial estradiol administration on recurrent insulin-induced hypoglycemia in ovariectomized female rats. Neuroendocrinology 96: 311-323. doi: 10.1159/000338407
|
| [11] |
Liu J, Bisschop PH, Eggels L, et al. (2013) Intrahypothalamic estradiol regulates glucose metabolism via the sympathetic nervous system in female rats. Diabetes 62: 435-443. doi: 10.2337/db12-0488
|
| [12] |
Lauber ME, Lichtensteiger W (1994) Pre- and postnatal ontogeny of aromatase cytochrome P450 messenger ribonucleic acid expression in the male rat brain studied by in situ hybridization. Endocrinology 135: 1661-1668. doi: 10.1210/endo.135.4.7925130
|
| [13] |
Roselli CE, Klosterman SA (1998) Sexual differentiation of aromatase activity in the rat brain: effects of perinatal steroid exposure. Endocrinology 139: 3193-3201. doi: 10.1210/endo.139.7.6101
|
| [14] |
Stanic D, Dubois S, Chua HK, et al. (2014) Characterization of aromatase expression in the adult male and female mouse brain. I. Coexistence with oestrogen receptors alpha and beta, and androgen receptors. PloS One 9: e90451. doi: 10.1371/journal.pone.0090451
|
| [15] | Uddin MM, Ibrahim MMH, Briski KP (2020) Ventromedial hypothalamic nucleus (VMN) neuroestradiol: sex mono-versus dimorphic control of glycogen metabolism and glucoregulatory transmitter signaling. Exp Biol 34: 1. |
| [16] |
Briski KP, Marshall ES, Sylvester PW (2001) Effects of estradiol on glucoprivic transactivation of catecholaminergic neurons in the female rat caudal brainstem. Neuroendocrinology 73: 369-377. doi: 10.1159/000054655
|
| [17] |
Nelson BS, Black KL, Daniel JM (2016) Circulating estradiol regulates brain-derived estradiol via actions at GnRH receptors to impact memory in ovariectomized rats. eNEURO 3. doi: 10.1523/ENEURO.0321-16.2016
|
| [18] |
Napit PR, Ali MH, Shakya M, et al. (2019) Hindbrain estrogen receptor regulation of counter-regulatory hormone secretion and ventromedial hypothalamic nucleus glycogen content and glucoregulatory transmitter signaling in hypoglycemic female rats. Neuroscience 411: 211-221. doi: 10.1016/j.neuroscience.2019.05.007
|
| [19] |
Mandal SK, Shrestha PK, Alenazi FSH, et al. (2018) Effects of estradiol on lactoprivic signaling of the hindbrain upon the contraregulatory hormonal response and metabolic neuropeptide synthesis in hypoglycemic female rats. Neuropeptides 70: 37-46. doi: 10.1016/j.npep.2018.05.004
|
| [20] |
Ibrahim MMH, Alhamami HN, Briski KP (2019) Norepinephrine regulation of ventromedial hypothalamic nucleus metabolic transmitter biomarker and astrocyte enzyme and receptor expression: impact of 5′-AMP-activated protein kinase. Brain Res 1711: 48-57. doi: 10.1016/j.brainres.2019.01.012
|
| [21] |
Kale AY, Paranjape SA, Briski KP (2006) I.c.v. administration of the nonsteroidal glucocorticoid receptor antagonist, CP4-72555, prevents exacerbated hypoglycemia during repeated insulin administration. Neuroscience 140: 555-565. doi: 10.1016/j.neuroscience.2006.02.041
|
| [22] |
Mahmood ASMH, Uddin MM, Mandal SK, et al. (2018) Sex differences in forebrain estrogen receptor regulation of hypoglycemic patterns of counter-regulatory hormone secretion and ventromedial hypothalamic nucleus gluco-regulatory neurotransmitter and astrocyte glycogen metabolic enzyme expression. Neuropeptides 72: 65-74. doi: 10.1016/j.npep.2018.10.003
|
| [23] |
Wagner CK, Morrell JI (1996) Distribution and steroid hormone regulation of aromatase mRNA expression in the forebrain of adult male and female rats: a cellular-level analysis using in situ hybridization. J Comp Neurol 370: 71-84. doi: 10.1002/(SICI)1096-9861(19960617)370:1<71::AID-CNE7>3.0.CO;2-I
|
| [24] |
Horvath TL, Wikler KC (1999) Aromatase in developing sensory systems of the rat brain. J Neuroendocrinol 11: 77-84. doi: 10.1046/j.1365-2826.1999.00285.x
|
| [25] |
Evrard H, Baillien M, Foidart A, et al. (2000) Localization and controls of aromatase in the quail spinal cord. J Comp Neurol 423: 552-564. doi: 10.1002/1096-9861(20000807)423:4<552::AID-CNE2>3.0.CO;2-S
|
| [26] |
Tran M, Kuhn JA, Bráz JM, et al. (2017) Neuronal aromatase expression in pain processing regions of the medullary and spinal cord dorsal horn. J Comp Neurol 525: 3414-3428. doi: 10.1002/cne.24269
|
| [27] |
Chan O, Zhu W, Ding Y, et al. (2006) Blockade of GABA (A) receptors in the ventromedial hypothalamus further stimulates glucagon and sympathoadrenal but not the hypothalamo-pituitary-adrenal response to hypoglycemia. Diabetes 55: 1080-1087. doi: 10.2337/diabetes.55.04.06.db05-0958
|
| [28] |
Routh VH, Hao L, Santiago AM, et al. (2014) Hypothalamic glucose sensing: making ends meet. Front Syst Neurosci 8: 236. doi: 10.3389/fnsys.2014.00236
|
| [29] |
Oomura Y, Ono H, Ooyama H, et al. (1969) Glucose and osmosensitive neurons of the rat hypothalamus. Nature 222: 282-284. doi: 10.1038/222282a0
|
| [30] |
Silver IA, Erecińska M (1998) Glucose-induced intracellular ion changes in sugar-sensitive hypothalamic neurons. J Neurophysiol 79: 1733-1745. doi: 10.1152/jn.1998.79.4.1733
|
| [31] |
Quenneville S, Labouèbe G, Basco D, et al. (2020) Hypoglycemia-sensing neurons of the ventromedial hypothalamus require AMPK-induced Txn2 expression but Are dispensable for physiological counterregulation. Diabetes 69: 2253-2266. doi: 10.2337/db20-0577
|
| [32] |
Ali MH, Napit PR, Mahmood ASMH, et al. (2019) Hindbrain estrogen receptor regulation of ventromedial hypothalamic glycogen metabolism and glucoregulatory transmitter expression in the hypoglycemic male rat. Neuroscience 409: 253-260. doi: 10.1016/j.neuroscience.2019.03.053
|

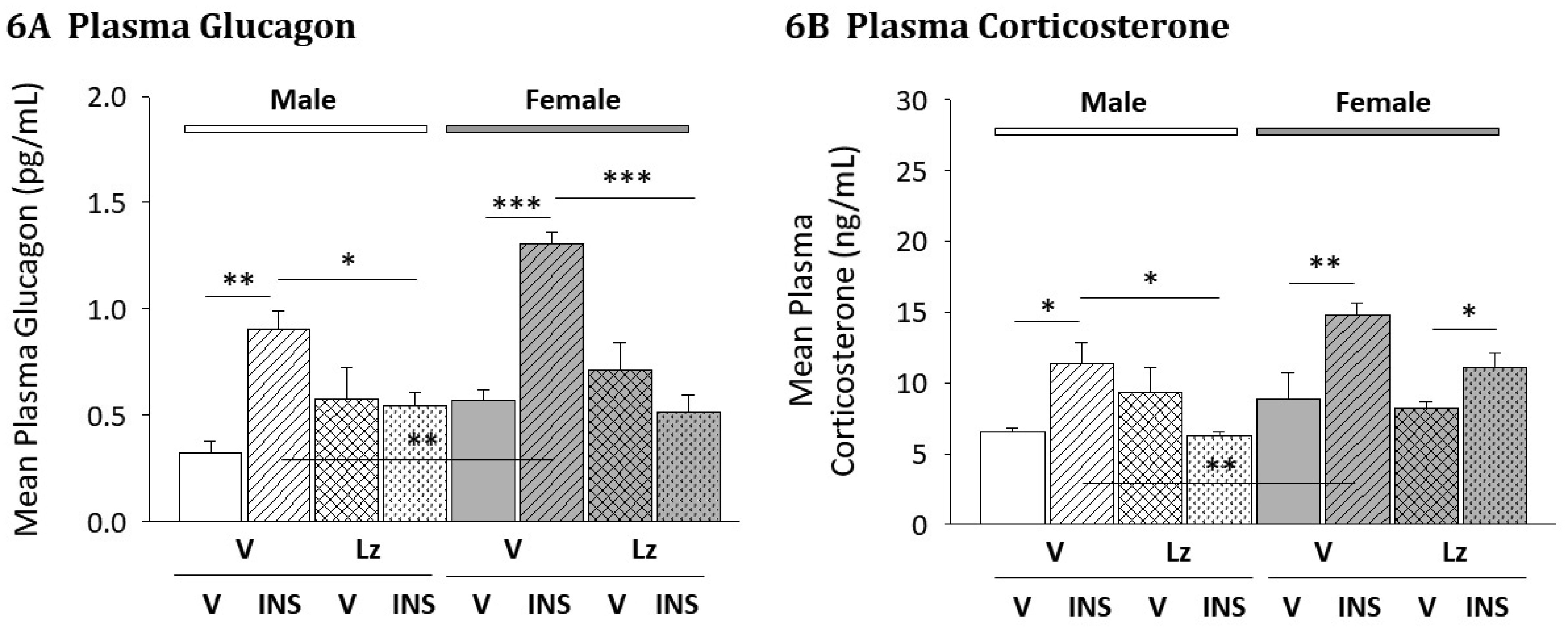
Figures(6) / Tables(1)

MD Main Uddin, Karen P Briski. Neuroestradiol regulation of ventromedial hypothalamic nucleus 5′-AMP-activated protein kinase activity and counterregulatory hormone secretion in hypoglycemic male versus female rats[J]. AIMS Neuroscience, 2021, 8(1): 133-147. doi: 10.3934/Neuroscience.2021006







 DownLoad:
DownLoad: