Citation: Marco Calabrò, Carmela Rinaldi, Giuseppe Santoro, Concetta Crisafulli. The biological pathways of Alzheimer disease: a review[J]. AIMS Neuroscience, 2021, 8(1): 86-132. doi: 10.3934/Neuroscience.2021005

| [1] | GBD 2015 Mortality and Causes of Death Collaborators (2016) Global, regional, and national life expectancy, all-cause mortality, and cause-specific mortality for 249 causes of death, 1980–2015: a systematic analysis for the Global Burden of Disease Study 2015. Lancet 388: 1459-1544. |
| [2] | APA (2020) 2020 Alzheimer's disease facts and figures. Alzheimers Dement in press. |
| [3] |
Brookmeyer R, Johnson E, Ziegler-Graham K, et al. (2007) Forecasting the global burden of Alzheimer's disease. Alzheimers Dement 3: 186-191. doi: 10.1016/j.jalz.2007.04.381

|
| [4] |
Prince M, Bryce R, Albanese E, et al. (2013) The global prevalence of dementia: a systematic review and metaanalysis. Alzheimers Dement 9: 63-75. e2. doi: 10.1016/j.jalz.2012.11.007
|
| [5] | Alzheimer's Association (2018) 2018 Alzheimer's disease facts and figures. Alzheimers Dement 14: 367-429. |
| [6] |
Perl DP (2010) Neuropathology of Alzheimer's disease. Mt Sinai J Med 77: 32-42. doi: 10.1002/msj.20157
|
| [7] | Kumar A, Tsao JW (2018) Alzheimer Disease StatPearls: Treasure Island (FL). |
| [8] |
Braak H, Braak E (1996) Development of Alzheimer-related neurofibrillary changes in the neocortex inversely recapitulates cortical myelogenesis. Acta Neuropathol 92: 197-201. doi: 10.1007/s004010050508
|
| [9] |
Cummings JL, Morstorf T, Zhong K (2014) Alzheimer's disease drug-development pipeline: few candidates, frequent failures. Alzheimers Res Ther 6: 37. doi: 10.1186/alzrt269
|
| [10] |
Hussein W, Saglik BN, Levent S, et al. (2018) Synthesis and biological evaluation of new cholinesterase inhibitors for Alzheimer's disease. Molecules 23: 2033. doi: 10.3390/molecules23082033
|
| [11] |
Leblhuber F, Steiner K, Schuetz B, et al. (2018) Probiotic supplementation in patients with Alzheimer's dementia—An explorative intervention study. Curr Alzheimer Res 15: 1106-1113. doi: 10.2174/1389200219666180813144834
|
| [12] |
Farlow MR, Salloway S, Tariot PN, et al. (2010) Effectiveness and tolerability of high-dose (23 mg/d) versus standard-dose (10 mg/d) donepezil in moderate to severe Alzheimer's disease: A 24-week, randomized, double-blind study. Clin Ther 32: 1234-1251. doi: 10.1016/j.clinthera.2010.06.019
|
| [13] |
Homma A, Atarashi H, Kubota N, et al. (2016) Efficacy and safety of sustained release donepezil high dose versus immediate release donepezil standard dose in Japanese patients with severe Alzheimer's disease: a randomized, double-blind trial. J Alzheimers Dis 52: 345-357. doi: 10.3233/JAD-151149
|
| [14] |
Winblad B, Kilander L, Eriksson S, et al. (2006) Donepezil in patients with severe Alzheimer's disease: double-blind, parallel-group, placebo-controlled study. Lancet 367: 1057-1065. doi: 10.1016/S0140-6736(06)68350-5
|
| [15] |
Feldman H, Gauthier S, Hecker J, et al. (2005) Efficacy and safety of donepezil in patients with more severe Alzheimer's disease: a subgroup analysis from a randomized, placebo-controlled trial. Int J Geriatr Psychiatry 20: 559-569. doi: 10.1002/gps.1325
|
| [16] |
Black SE, Doody R, Li H, et al. (2007) Donepezil preserves cognition and global function in patients with severe Alzheimer disease. Neurology 69: 459-469. doi: 10.1212/01.wnl.0000266627.96040.5a
|
| [17] |
Howard R, McShane R, Lindesay J, et al. (2015) Nursing home placement in the donepezil and memantine in moderate to severe Alzheimer's disease (DOMINO-AD) trial: secondary and post-hoc analyses. Lancet Neurol 14: 1171-1181. doi: 10.1016/S1474-4422(15)00258-6
|
| [18] |
Bond M, Rogers G, Peters J, et al. (2012) The effectiveness and cost-effectiveness of donepezil, galantamine, rivastigmine and memantine for the treatment of Alzheimer's disease (review of Technology Appraisal No. 111): a systematic review and economic model. Health Technol Assess 16: 1-470. doi: 10.3310/hta16210
|
| [19] |
Zhang N, Wei C, Du H, et al. (2015) The effect of memantine on cognitive function and behavioral and psychological symptoms in mild-to-moderate Alzheimer's disease patients. Dement Geriatr Cogn Disord 40: 85-93. doi: 10.1159/000430808
|
| [20] |
Molinuevo JL, Berthier ML, Rami L (2011) Donepezil provides greater benefits in mild compared to moderate Alzheimer's disease: implications for early diagnosis and treatment. Arch Gerontol Geriatr 52: 18-22. doi: 10.1016/j.archger.2009.11.004
|
| [21] |
Takeda A, Loveman E, Clegg A, et al. (2006) A systematic review of the clinical effectiveness of donepezil, rivastigmine and galantamine on cognition, quality of life and adverse events in Alzheimer's disease. Int J Geriatr Psychiatry 21: 17-28. doi: 10.1002/gps.1402
|
| [22] |
Lam B, Masellis M, Freedman M, et al. (2013) Clinical, imaging, and pathological heterogeneity of the Alzheimer's disease syndrome. Alzheimers Res Ther 5: 1-14. doi: 10.1186/alzrt155
|
| [23] |
Jack CR, Albert MS, Knopman DS, et al. (2011) Introduction to the recommendations from the National Institute on Aging-Alzheimer's Association workgroups on diagnostic guidelines for Alzheimer's disease. Alzheimers Dement 7: 257-262. doi: 10.1016/j.jalz.2011.03.004
|
| [24] |
Albert MS, DeKosky ST, Dickson D, et al. (2011) The diagnosis of mild cognitive impairment due to Alzheimer's disease: recommendations from the National Institute on Aging-Alzheimer's Association workgroups on diagnostic guidelines for Alzheimer's disease. Alzheimers Dement 7: 270-279. doi: 10.1016/j.jalz.2011.03.008
|
| [25] |
McKhann GM, Knopman DS, Chertkow H, et al. (2011) The diagnosis of dementia due to Alzheimer's disease: recommendations from the National Institute on Aging-Alzheimer's Association workgroups on diagnostic guidelines for Alzheimer's disease. Alzheimers Dement 7: 263-269. doi: 10.1016/j.jalz.2011.03.005
|
| [26] |
Beason-Held LL, Goh JO, An Y, et al. (2013) Changes in brain function occur years before the onset of cognitive impairment. J Neurosci 33: 18008-18014. doi: 10.1523/JNEUROSCI.1402-13.2013
|
| [27] |
O'Brien RJ, Wong PC (2011) Amyloid precursor protein processing and Alzheimer's disease. Annu Rev Neurosci 34: 185-204. doi: 10.1146/annurev-neuro-061010-113613
|
| [28] |
Wilkins HM, Swerdlow RH (2017) Amyloid precursor protein processing and bioenergetics. Brain Res Bull 133: 71-79. doi: 10.1016/j.brainresbull.2016.08.009
|
| [29] |
Vassar R, Kovacs DM, Yan R, et al. (2009) The beta-secretase enzyme BACE in health and Alzheimer's disease: regulation, cell biology, function, and therapeutic potential. J Neurosci 29: 12787-12794. doi: 10.1523/JNEUROSCI.3657-09.2009
|
| [30] |
De Strooper B, Vassar R, Golde T (2010) The secretases: enzymes with therapeutic potential in Alzheimer disease. Nat Rev Neurol 6: 99-107. doi: 10.1038/nrneurol.2009.218
|
| [31] |
Walsh DM, Klyubin I, Fadeeva JV, et al. (2002) Naturally secreted oligomers of amyloid beta protein potently inhibit hippocampal long-term potentiation in vivo. Nature 416: 535-539. doi: 10.1038/416535a
|
| [32] |
Chen YY, Wang MC, Wang YN, et al. (2020) Redox signaling and Alzheimer's disease: from pathomechanism insights to biomarker discovery and therapy strategy. Biomark Res 8: 42. doi: 10.1186/s40364-020-00218-z
|
| [33] |
Alonso AC, Zaidi T, Grundke-Iqbal I, et al. (1994) Role of abnormally phosphorylated tau in the breakdown of microtubules in Alzheimer disease. Proc Natl Acad Sci U S A 91: 5562-5566. doi: 10.1073/pnas.91.12.5562
|
| [34] |
Alonso AC, Grundke-Iqbal I, Iqbal K (1996) Alzheimer's disease hyperphosphorylated tau sequesters normal tau into tangles of filaments and disassembles microtubules. Nat Med 2: 783-787. doi: 10.1038/nm0796-783
|
| [35] |
Braak H, Alafuzoff I, Arzberger T, et al. (2006) Staging of Alzheimer disease-associated neurofibrillary pathology using paraffin sections and immunocytochemistry. Acta Neuropathol 112: 389-404. doi: 10.1007/s00401-006-0127-z
|
| [36] |
Morrison BM, Hof PR, Morrison JH (1998) Determinants of neuronal vulnerability in neurodegenerative diseases. Ann Neurol 44: S32-44. doi: 10.1002/ana.410440706
|
| [37] |
Braak H, Braak E (1985) On areas of transition between entorhinal allocortex and temporal isocortex in the human brain. Normal morphology and lamina-specific pathology in Alzheimer's disease. Acta Neuropathol 68: 325-332. doi: 10.1007/BF00690836
|
| [38] |
Mesulam MM (1999) Neuroplasticity failure in Alzheimer's disease: bridging the gap between plaques and tangles. Neuron 24: 521-529. doi: 10.1016/S0896-6273(00)81109-5
|
| [39] |
Serrano-Pozo A, Frosch MP, Masliah E, et al. (2011) Neuropathological alterations in Alzheimer disease. Cold Spring Harb Perspect Med 1: a006189. doi: 10.1101/cshperspect.a006189
|
| [40] |
Goldman JS, Hahn SE, Catania JW, et al. (2011) Genetic counseling and testing for Alzheimer disease: joint practice guidelines of the American College of Medical Genetics and the National Society of Genetic Counselors. Genet Med 13: 597-605. doi: 10.1097/GIM.0b013e31821d69b8
|
| [41] |
Bekris LM, Yu CE, Bird TD, et al. (2010) Genetics of Alzheimer disease. J Geriatr Psychiatry Neurol 23: 213-227. doi: 10.1177/0891988710383571
|
| [42] |
Brickell KL, Steinbart EJ, Rumbaugh M, et al. (2006) Early-onset Alzheimer disease in families with late-onset Alzheimer disease: a potential important subtype of familial Alzheimer disease. Arch Neurol 63: 1307-1311. doi: 10.1001/archneur.63.9.1307
|
| [43] |
Bird TD (2008) Genetic aspects of Alzheimer disease. Genet Med 10: 231-239. doi: 10.1097/GIM.0b013e31816b64dc
|
| [44] |
Serretti A, Artioli P, Quartesan R, et al. (2005) Genes involved in Alzheimer's disease, a survey of possible candidates. J Alzheimers Dis 7: 331-353. doi: 10.3233/JAD-2005-7410
|
| [45] |
Mahley RW (1988) Apolipoprotein E: cholesterol transport protein with expanding role in cell biology. Science 240: 622-630. doi: 10.1126/science.3283935
|
| [46] |
Puglielli L, Tanzi RE, Kovacs DM (2003) Alzheimer's disease: the cholesterol connection. Nat Neurosci 6: 345-351. doi: 10.1038/nn0403-345
|
| [47] |
Harold D, Abraham R, Hollingworth P, et al. (2009) Genome-wide association study identifies variants at CLU and PICALM associated with Alzheimer's disease. Nat Genet 41: 1088-1093. doi: 10.1038/ng.440
|
| [48] |
Lambert JC, Heath S, Even G, et al. (2009) Genome-wide association study identifies variants at CLU and CR1 associated with Alzheimer's disease. Nat Genet 41: 1094-1099. doi: 10.1038/ng.439
|
| [49] |
Saunders AM, Strittmatter WJ, Schmechel D, et al. (1993) Association of apolipoprotein E allele epsilon 4 with late-onset familial and sporadic Alzheimer's disease. Neurology 43: 1467-1472. doi: 10.1212/WNL.43.8.1467
|
| [50] |
Farrer LA, Cupples LA, Haines JL, et al. (1997) Effects of age, sex, and ethnicity on the association between apolipoprotein E genotype and Alzheimer disease. A meta-analysis. APOE and Alzheimer Disease Meta Analysis Consortium. JAMA 278: 1349-1356. doi: 10.1001/jama.1997.03550160069041
|
| [51] |
Zlokovic BV (2013) Cerebrovascular effects of apolipoprotein E: implications for Alzheimer disease. JAMA Neurol 70: 440-444. doi: 10.1001/jamaneurol.2013.2152
|
| [52] |
Ghebremedhin E, Schultz C, Braak E, et al. (1998) High frequency of apolipoprotein E epsilon4 allele in young individuals with very mild Alzheimer's disease-related neurofibrillary changes. Exp Neurol 153: 152-155. doi: 10.1006/exnr.1998.6860
|
| [53] |
Shi Y, Yamada K, Liddelow SA, et al. (2017) ApoE4 markedly exacerbates tau-mediated neurodegeneration in a mouse model of tauopathy. Nature 549: 523-527. doi: 10.1038/nature24016
|
| [54] |
Ulrich JD, Ulland TK, Mahan TE, et al. (2018) ApoE facilitates the microglial response to amyloid plaque pathology. J Exp Med 215: 1047-1058. doi: 10.1084/jem.20171265
|
| [55] |
Mahley RW (2016) Central nervous system lipoproteins: ApoE and regulation of cholesterol metabolism. Arterioscler Thromb Vasc Biol 36: 1305-1315. doi: 10.1161/ATVBAHA.116.307023
|
| [56] |
Yamazaki Y, Zhao N, Caulfield TR, et al. (2019) Apolipoprotein E and Alzheimer disease: pathobiology and targeting strategies. Nat Rev Neurol 15: 501-518. doi: 10.1038/s41582-019-0228-7
|
| [57] |
Verghese PB, Castellano JM, Garai K, et al. (2013) ApoE influences amyloid-beta (Abeta) clearance despite minimal apoE/Abeta association in physiological conditions. Proc Natl Acad Sci U S A 110: E1807-1816. doi: 10.1073/pnas.1220484110
|
| [58] |
Huang Y, Mahley RW (2014) Apolipoprotein E: structure and function in lipid metabolism, neurobiology, and Alzheimer's diseases. Neurobiol Dis 72: 3-12. doi: 10.1016/j.nbd.2014.08.025
|
| [59] |
Miyata M, Smith JD (1996) Apolipoprotein E allele-specific antioxidant activity and effects on cytotoxicity by oxidative insults and beta-amyloid peptides. Nat Genet 14: 55-61. doi: 10.1038/ng0996-55
|
| [60] | Montine KS, Olson SJ, Amarnath V, et al. (1997) Immunohistochemical detection of 4-hydroxy-2-nonenal adducts in Alzheimer's disease is associated with inheritance of APOE4. Am J Pathol 150: 437-443. |
| [61] |
Ramassamy C, Averill D, Beffert U, et al. (1999) Oxidative damage and protection by antioxidants in the frontal cortex of Alzheimer's disease is related to the apolipoprotein E genotype. Free Radic Biol Med 27: 544-553. doi: 10.1016/S0891-5849(99)00102-1
|
| [62] |
Chen Y, Zhang J, Zhang B, et al. (2016) Targeting insulin signaling for the treatment of Alzheimer's disease. Curr Top Med Chem 16: 485-492. doi: 10.2174/1568026615666150813142423
|
| [63] |
Mahley RW, Huang Y (2012) Apolipoprotein e sets the stage: response to injury triggers neuropathology. Neuron 76: 871-885. doi: 10.1016/j.neuron.2012.11.020
|
| [64] |
Zhong N, Weisgraber KH (2009) Understanding the association of apolipoprotein E4 with Alzheimer disease: clues from its structure. J Biol Chem 284: 6027-6031. doi: 10.1074/jbc.R800009200
|
| [65] |
Reiman EM, Caselli RJ, Chen K, et al. (2001) Declining brain activity in cognitively normal apolipoprotein E epsilon 4 heterozygotes: a foundation for using positron emission tomography to efficiently test treatments to prevent Alzheimer's disease. Proc Natl Acad Sci U S A 98: 3334-3339. doi: 10.1073/pnas.061509598
|
| [66] |
Reiman EM, Chen K, Alexander GE, et al. (2004) Functional brain abnormalities in young adults at genetic risk for late-onset Alzheimer's dementia. Proc Natl Acad Sci U S A 101: 284-289. doi: 10.1073/pnas.2635903100
|
| [67] |
Skotte N (2010) Genome-wide association studies identify new interesting loci for late-onset Alzheimer's disease. Clin Genet 77: 330-332. doi: 10.1111/j.1399-0004.2009.01366_3.x
|
| [68] |
Chouraki V, Seshadri S (2014) Genetics of Alzheimer's disease. Adv Genet 87: 245-294. doi: 10.1016/B978-0-12-800149-3.00005-6
|
| [69] |
Bertram L, McQueen MB, Mullin K, et al. (2007) Systematic meta-analyses of Alzheimer disease genetic association studies: the AlzGene database. Nat Genet 39: 17-23. doi: 10.1038/ng1934
|
| [70] |
Goldgaber D, Lerman MI, McBride OW, et al. (1987) Characterization and chromosomal localization of a cDNA encoding brain amyloid of Alzheimer's disease. Science 235: 877-880. doi: 10.1126/science.3810169
|
| [71] |
Kang J, Lemaire HG, Unterbeck A, et al. (1987) The precursor of Alzheimer's disease amyloid A4 protein resembles a cell-surface receptor. Nature 325: 733-736. doi: 10.1038/325733a0
|
| [72] |
Robakis NK, Wisniewski HM, Jenkins EC, et al. (1987) Chromosome 21q21 sublocalisation of gene encoding beta-amyloid peptide in cerebral vessels and neuritic (senile) plaques of people with Alzheimer disease and Down syndrome. Lancet 1: 384-385. doi: 10.1016/S0140-6736(87)91754-5
|
| [73] |
Tanzi RE, Gusella JF, Watkins PC, et al. (1987) Amyloid beta protein gene: cDNA, mRNA distribution, and genetic linkage near the Alzheimer locus. Science 235: 880-884. doi: 10.1126/science.2949367
|
| [74] |
Sherrington R, Rogaev EI, Liang Y, et al. (1995) Cloning of a gene bearing missense mutations in early-onset familial Alzheimer's disease. Nature 375: 754-760. doi: 10.1038/375754a0
|
| [75] |
Levy-Lahad E, Wasco W, Poorkaj P, et al. (1995) Candidate gene for the chromosome 1 familial Alzheimer's disease locus. Science 269: 973-977. doi: 10.1126/science.7638622
|
| [76] |
Zhao QF, Yu JT, Tan MS, et al. (2015) ABCA7 in Alzheimer's Disease. Mol Neurobiol 51: 1008-1016. doi: 10.1007/s12035-014-8759-9
|
| [77] |
Hollingworth P, Harold D, Sims R, et al. (2011) Common variants at ABCA7, MS4A6A/MS4A4E, EPHA1, CD33 and CD2AP are associated with Alzheimer's disease. Nat Genet 43: 429-435. doi: 10.1038/ng.803
|
| [78] |
Marcello E, Saraceno C, Musardo S, et al. (2013) Endocytosis of synaptic ADAM10 in neuronal plasticity and Alzheimer's disease. J Clin Invest 123: 2523-2538. doi: 10.1172/JCI65401
|
| [79] |
Corder EH, Saunders AM, Risch NJ, et al. (1994) Protective effect of apolipoprotein E type 2 allele for late onset Alzheimer disease. Nat Genet 7: 180-184. doi: 10.1038/ng0694-180
|
| [80] |
Royston MC, Mann D, Pickering-Brown S, et al. (1994) Apolipoprotein E epsilon 2 allele promotes longevity and protects patients with Down's syndrome from dementia. Neuroreport 5: 2583-2585. doi: 10.1097/00001756-199412000-00044
|
| [81] |
Coimbra JRM, Marques DFF, Baptista SJ, et al. (2018) Highlights in BACE1 inhibitors for Alzheimer's disease treatment. Front Chem 6: 178. doi: 10.3389/fchem.2018.00178
|
| [82] |
Holsinger RM, Goense N, Bohorquez J, et al. (2013) Splice variants of the Alzheimer's disease beta-secretase, BACE1. Neurogenetics 14: 1-9. doi: 10.1007/s10048-012-0348-3
|
| [83] |
Tan MS, Yu JT, Tan L (2013) Bridging integrator 1 (BIN1): form, function, and Alzheimer's disease. Trends Mol Med 19: 594-603. doi: 10.1016/j.molmed.2013.06.004
|
| [84] |
Seshadri S, Fitzpatrick AL, Ikram MA, et al. (2010) Genome-wide analysis of genetic loci associated with Alzheimer disease. JAMA 303: 1832-1840. doi: 10.1001/jama.2010.574
|
| [85] |
Liu G, Zhang S, Cai Z, et al. (2013) BIN1 gene rs744373 polymorphism contributes to Alzheimer's disease in East Asian population. Neurosci Lett 544: 47-51. doi: 10.1016/j.neulet.2013.02.075
|
| [86] |
Miyashita A, Koike A, Jun G, et al. (2013) SORL1 is genetically associated with late-onset Alzheimer's disease in Japanese, Koreans and Caucasians. PLoS One 8: e58618. doi: 10.1371/journal.pone.0058618
|
| [87] |
Harms M, Benitez BA, Cairns N, et al. (2013) C9orf72 hexanucleotide repeat expansions in clinical Alzheimer disease. JAMA Neurol 70: 736-741. doi: 10.1001/2013.jamaneurol.537
|
| [88] |
Jiang T, Yu JT, Hu N, et al. (2014) CD33 in Alzheimer's disease. Mol Neurobiol 49: 529-535. doi: 10.1007/s12035-013-8536-1
|
| [89] |
Deng YL, Liu LH, Wang Y, et al. (2012) The prevalence of CD33 and MS4A6A variant in Chinese Han population with Alzheimer's disease. Hum Genet 131: 1245-1249. doi: 10.1007/s00439-012-1154-6
|
| [90] |
Tan L, Yu JT, Zhang W, et al. (2013) Association of GWAS-linked loci with late-onset Alzheimer's disease in a northern Han Chinese population. Alzheimers Dement 9: 546-553. doi: 10.1016/j.jalz.2012.08.007
|
| [91] |
Liu G, Wang H, Liu J, et al. (2014) The CLU gene rs11136000 variant is significantly associated with Alzheimer's disease in Caucasian and Asian populations. Neuromolecular Med 16: 52-60. doi: 10.1007/s12017-013-8250-1
|
| [92] |
Komatsu M, Shibata N, Kuerban B, et al. (2011) Genetic association between clusterin polymorphisms and Alzheimer's disease in a Japanese population. Psychogeriatrics 11: 14-18. doi: 10.1111/j.1479-8301.2010.00346.x
|
| [93] |
Yu JT, Ma XY, Wang YL, et al. (2013) Genetic variation in clusterin gene and Alzheimer's disease risk in Han Chinese. Neurobiol Aging 34: 1921. e17-23. doi: 10.1016/j.neurobiolaging.2013.01.010
|
| [94] |
Jin C, Li W, Yuan J, et al. (2012) Association of the CR1 polymorphism with late-onset Alzheimer's disease in Chinese Han populations: a meta-analysis. Neurosci Lett 527: 46-49. doi: 10.1016/j.neulet.2012.08.032
|
| [95] |
Crehan H, Holton P, Wray S, et al. (2012) Complement receptor 1 (CR1) and Alzheimer's disease. Immunobiology 217: 244-250. doi: 10.1016/j.imbio.2011.07.017
|
| [96] |
Nacmias B, Piaceri I, Bagnoli S, et al. (2014) Genetics of Alzheimer's disease and frontotemporal dementia. Curr Mol Med 14: 993-1000. doi: 10.2174/1566524014666141010152143
|
| [97] |
Perry DC, Lehmann M, Yokoyama JS, et al. (2013) Progranulin mutations as risk factors for Alzheimer disease. JAMA Neurol 70: 774-778. doi: 10.1001/2013.jamaneurol.393
|
| [98] |
Li HL, Lu SJ, Sun YM, et al. (2013) The LRRK2 R1628P variant plays a protective role in Han Chinese population with Alzheimer's disease. CNS Neurosci Ther 19: 207-215. doi: 10.1111/cns.12062
|
| [99] |
Zhao Y, Ho P, Yih Y, et al. (2011) LRRK2 variant associated with Alzheimer's disease. Neurobiol Aging 32: 1990-1993. doi: 10.1016/j.neurobiolaging.2009.11.019
|
| [100] |
Parikh I, Fardo DW, Estus S (2014) Genetics of PICALM expression and Alzheimer's disease. PLoS One 9: e91242. doi: 10.1371/journal.pone.0091242
|
| [101] |
Chung SJ, Lee JH, Kim SY, et al. (2013) Association of GWAS top hits with late-onset Alzheimer disease in Korean population. Alzheimer Dis Assoc Disord 27: 250-257. doi: 10.1097/WAD.0b013e31826d7281
|
| [102] |
Liu G, Zhang S, Cai Z, et al. (2013) PICALM gene rs3851179 polymorphism contributes to Alzheimer's disease in an Asian population. Neuromolecular Med 15: 384-388. doi: 10.1007/s12017-013-8225-2
|
| [103] |
Wang Q, Tian Q, Song X, et al. (2016) SNCA gene polymorphism may contribute to an increased risk of Alzheimer's disease. J Clin Lab Anal 30: 1092-1099. doi: 10.1002/jcla.21986
|
| [104] |
Linnertz C, Lutz MW, Ervin JF, et al. (2014) The genetic contributions of SNCA and LRRK2 genes to Lewy body pathology in Alzheimer's disease. Hum Mol Genet 23: 4814-4821. doi: 10.1093/hmg/ddu196
|
| [105] |
Vardarajan BN, Zhang Y, Lee JH, et al. (2015) Coding mutations in SORL1 and Alzheimer disease. Ann Neurol 77: 215-227. doi: 10.1002/ana.24305
|
| [106] |
Yin RH, Yu JT, Tan L (2015) The role of SORL1 in Alzheimer's disease. Mol Neurobiol 51: 909-918. doi: 10.1007/s12035-014-8742-5
|
| [107] |
Rogaeva E, Meng Y, Lee JH, et al. (2007) The neuronal sortilin-related receptor SORL1 is genetically associated with Alzheimer disease. Nat Genet 39: 168-177. doi: 10.1038/ng1943
|
| [108] |
Lambert JC, Ibrahim-Verbaas CA, Harold D, et al. (2013) Meta-analysis of 74,046 individuals identifies 11 new susceptibility loci for Alzheimer's disease. Nat Genet 45: 1452-1458. doi: 10.1038/ng.2802
|
| [109] |
Allen M, Kachadoorian M, Quicksall Z, et al. (2014) Association of MAPT haplotypes with Alzheimer's disease risk and MAPT brain gene expression levels. Alzheimers Res Ther 6: 39. doi: 10.1186/alzrt268
|
| [110] |
Zhang N, Yu JT, Yang Y, et al. (2011) Association analysis of GSK3B and MAPT polymorphisms with Alzheimer's disease in Han Chinese. Brain Res 1391: 147-153. doi: 10.1016/j.brainres.2011.03.052
|
| [111] |
Bloom GS (2014) Amyloid-beta and tau: the trigger and bullet in Alzheimer disease pathogenesis. JAMA Neurol 71: 505-508. doi: 10.1001/jamaneurol.2013.5847
|
| [112] |
James BD, Wilson RS, Boyle PA, et al. (2016) TDP-43 stage, mixed pathologies, and clinical Alzheimer's-type dementia. Brain 139: 2983-2993. doi: 10.1093/brain/aww224
|
| [113] |
Brouwers N, Bettens K, Gijselinck I, et al. (2010) Contribution of TARDBP to Alzheimer's disease genetic etiology. J Alzheimers Dis 21: 423-430. doi: 10.3233/JAD-2010-100198
|
| [114] |
Guerreiro R, Wojtas A, Bras J, et al. (2013) TREM2 variants in Alzheimer's disease. N Engl J Med 368: 117-127. doi: 10.1056/NEJMoa1211851
|
| [115] |
Lu Y, Liu W, Wang X (2015) TREM2 variants and risk of Alzheimer's disease: a meta-analysis. Neurol Sci 36: 1881-1888. doi: 10.1007/s10072-015-2274-2
|
| [116] |
Naj AC, Jun G, Beecham GW, et al. (2011) Common variants at MS4A4/MS4A6E, CD2AP, CD33 and EPHA1 are associated with late-onset Alzheimer's disease. Nat Genet 43: 436-441. doi: 10.1038/ng.801
|
| [117] |
Wijsman EM, Pankratz ND, Choi Y, et al. (2011) Genome-wide association of familial late-onset Alzheimer's disease replicates BIN1 and CLU and nominates CUGBP2 in interaction with APOE. PLoS Genet 7: e1001308. doi: 10.1371/journal.pgen.1001308
|
| [118] |
Boada M, Antunez C, Ramirez-Lorca R, et al. (2014) ATP5H/KCTD2 locus is associated with Alzheimer's disease risk. Mol Psychiatry 19: 682-687. doi: 10.1038/mp.2013.86
|
| [119] |
Beecham GW, Naj AC, Gilbert JR, et al. (2010) PCDH11X variation is not associated with late-onset Alzheimer disease susceptibility. Psychiatr Genet 20: 321-324. doi: 10.1097/YPG.0b013e32833b635d
|
| [120] |
Miar A, Alvarez V, Corao AI, et al. (2011) Lack of association between protocadherin 11-X/Y (PCDH11X and PCDH11Y) polymorphisms and late onset Alzheimer's disease. Brain Res 1383: 252-256. doi: 10.1016/j.brainres.2011.01.054
|
| [121] |
Jiang T, Yu JT, Wang YL, et al. (2014) The genetic variation of ARRB2 is associated with late-onset Alzheimer's disease in Han Chinese. Curr Alzheimer Res 11: 408-412. doi: 10.2174/1567205011666140317095014
|
| [122] |
Gorski DH, Walsh K (2003) Control of vascular cell differentiation by homeobox transcription factors. Trends Cardiovasc Med 13: 213-220. doi: 10.1016/S1050-1738(03)00081-1
|
| [123] |
Rovelet-Lecrux A, Legallic S, Wallon D, et al. (2012) A genome-wide study reveals rare CNVs exclusive to extreme phenotypes of Alzheimer disease. Eur J Hum Genet 20: 613-617. doi: 10.1038/ejhg.2011.225
|
| [124] |
Wu Z, Guo H, Chow N, et al. (2005) Role of the MEOX2 homeobox gene in neurovascular dysfunction in Alzheimer disease. Nat Med 11: 959-965. doi: 10.1038/nm1287
|
| [125] |
Mielke MM, Vemuri P, Rocca WA (2014) Clinical epidemiology of Alzheimer's disease: assessing sex and gender differences. Clin Epidemiol 6: 37-48. doi: 10.2147/CLEP.S37929
|
| [126] |
Fjell AM, McEvoy L, Holland D, et al. (2014) What is normal in normal aging? Effects of aging, amyloid and Alzheimer's disease on the cerebral cortex and the hippocampus. Prog Neurobiol 117: 20-40. doi: 10.1016/j.pneurobio.2014.02.004
|
| [127] | Lee JY, Chang SM, Jang HS, et al. (2008) Illiteracy and the incidence of Alzheimer's disease in the Yonchon County survey, Korea. Int Psychogeriatr 20: 976-985. |
| [128] |
Durazzo TC, Mattsson N, Weiner MW, et al. (2014) Smoking and increased Alzheimer's disease risk: a review of potential mechanisms. Alzheimers Dement 10: S122-145. doi: 10.1016/j.jalz.2014.04.009
|
| [129] |
Barnard ND, Bush AI, Ceccarelli A, et al. (2014) Dietary and lifestyle guidelines for the prevention of Alzheimer's disease. Neurobiol Aging 35: S74-78. doi: 10.1016/j.neurobiolaging.2014.03.033
|
| [130] |
Kawahara M, Kato-Negishi M, Tanaka K (2017) Cross talk between neurometals and amyloidogenic proteins at the synapse and the pathogenesis of neurodegenerative diseases. Metallomics 9: 619-633. doi: 10.1039/C7MT00046D
|
| [131] |
Zheng W, Aschner M, Ghersi-Egea JF (2003) Brain barrier systems: a new frontier in metal neurotoxicological research. Toxicol Appl Pharmacol 192: 1-11. doi: 10.1016/S0041-008X(03)00251-5
|
| [132] |
Heneka MT, Carson MJ, El Khoury J, et al. (2015) Neuroinflammation in Alzheimer's disease. Lancet Neurol 14: 388-405. doi: 10.1016/S1474-4422(15)70016-5
|
| [133] |
Wang X, Wang W, Li L, et al. (2014) Oxidative stress and mitochondrial dysfunction in Alzheimer's disease. Biochim Biophys Acta 1842: 1240-1247. doi: 10.1016/j.bbadis.2013.10.015
|
| [134] |
Chen Z, Zhong C (2014) Oxidative stress in Alzheimer's disease. Neurosci Bull 30: 271-281. doi: 10.1007/s12264-013-1423-y
|
| [135] |
DuBoff B, Feany M, Gotz J (2013) Why size matters-balancing mitochondrial dynamics in Alzheimer's disease. Trends Neurosci 36: 325-335. doi: 10.1016/j.tins.2013.03.002
|
| [136] |
Dik MG, Jonker C, Comijs HC, et al. (2007) Contribution of metabolic syndrome components to cognition in older individuals. Diabetes Care 30: 2655-2660. doi: 10.2337/dc06-1190
|
| [137] |
Campos-Pena V, Toral-Rios D, Becerril-Perez F, et al. (2017) Metabolic syndrome as a risk factor for Alzheimer's disease: is abeta a crucial factor in both pathologies? Antioxid Redox Signal 26: 542-560. doi: 10.1089/ars.2016.6768
|
| [138] |
Whyte LS, Lau AA, Hemsley KM, et al. (2017) Endo-lysosomal and autophagic dysfunction: a driving factor in Alzheimer's disease? J Neurochem 140: 703-717. doi: 10.1111/jnc.13935
|
| [139] |
Ferreira ST, Clarke JR, Bomfim TR, et al. (2014) Inflammation, defective insulin signaling, and neuronal dysfunction in Alzheimer's disease. Alzheimers Dement 10: S76-83. doi: 10.1016/j.jalz.2013.12.010
|
| [140] |
Kinney JW, Bemiller SM, Murtishaw AS, et al. (2018) Inflammation as a central mechanism in Alzheimer's disease. Alzheimers Dement 4: 575-590. doi: 10.1016/j.trci.2018.06.014
|
| [141] | Rubio-Perez JM, Morillas-Ruiz JM (2012) A review: inflammatory process in Alzheimer's disease, role of cytokines. Sci World J 2012: 756357. |
| [142] |
Mengel-From J, Christensen K, McGue M, et al. (2011) Genetic variations in the CLU and PICALM genes are associated with cognitive function in the oldest old. Neurobiol Aging 32: 554. e7-11. doi: 10.1016/j.neurobiolaging.2010.07.016
|
| [143] |
Jonsson T, Stefansson H, Steinberg S, et al. (2013) Variant of TREM2 associated with the risk of Alzheimer's disease. N Engl J Med 368: 107-116. doi: 10.1056/NEJMoa1211103
|
| [144] |
Cunningham C (2013) Microglia and neurodegeneration: the role of systemic inflammation. Glia 61: 71-90. doi: 10.1002/glia.22350
|
| [145] |
Bodea LG, Wang Y, Linnartz-Gerlach B, et al. (2014) Neurodegeneration by activation of the microglial complement-phagosome pathway. J Neurosci 34: 8546-8556. doi: 10.1523/JNEUROSCI.5002-13.2014
|
| [146] |
Mhatre SD, Tsai CA, Rubin AJ, et al. (2015) Microglial malfunction: the third rail in the development of Alzheimer's disease. Trends Neurosci 38: 621-636. doi: 10.1016/j.tins.2015.08.006
|
| [147] |
Zotova E, Nicoll JA, Kalaria R, et al. (2010) Inflammation in Alzheimer's disease: relevance to pathogenesis and therapy. Alzheimers Res Ther 2: 1. doi: 10.1186/alzrt24
|
| [148] |
Musicco C, Capelli V, Pesce V, et al. (2009) Accumulation of overoxidized peroxiredoxin III in aged rat liver mitochondria. Biochim Biophys Acta 1787: 890-896. doi: 10.1016/j.bbabio.2009.03.002
|
| [149] |
Sutinen EM, Pirttila T, Anderson G, et al. (2012) Pro-inflammatory interleukin-18 increases Alzheimer's disease-associated amyloid-beta production in human neuron-like cells. J Neuroinflammation 9: 199. doi: 10.1186/1742-2094-9-199
|
| [150] |
Sutinen EM, Korolainen MA, Hayrinen J, et al. (2014) Interleukin-18 alters protein expressions of neurodegenerative diseases-linked proteins in human SH-SY5Y neuron-like cells. Front Cell Neurosci 8: 214. doi: 10.3389/fncel.2014.00214
|
| [151] | Oakley R, Tharakan B (2014) Vascular hyperpermeability and aging. Aging Dis 5: 114-125. |
| [152] |
McColl BW, Rose N, Robson FH, et al. (2010) Increased brain microvascular MMP-9 and incidence of haemorrhagic transformation in obese mice after experimental stroke. J Cereb Blood Flow Metab 30: 267-272. doi: 10.1038/jcbfm.2009.217
|
| [153] |
Ojala JO, Sutinen EM, Salminen A, et al. (2008) Interleukin-18 increases expression of kinases involved in tau phosphorylation in SH-SY5Y neuroblastoma cells. J Neuroimmunol 205: 86-93. doi: 10.1016/j.jneuroim.2008.09.012
|
| [154] |
Alvarez A, Toro R, Caceres A, et al. (1999) Inhibition of tau phosphorylating protein kinase cdk5 prevents beta-amyloid-induced neuronal death. FEBS Lett 459: 421-426. doi: 10.1016/S0014-5793(99)01279-X
|
| [155] |
Alvarez A, Munoz JP, Maccioni RB (2001) A Cdk5-p35 stable complex is involved in the beta-amyloid-induced deregulation of Cdk5 activity in hippocampal neurons. Exp Cell Res 264: 266-274. doi: 10.1006/excr.2001.5152
|
| [156] |
Seo J, Kritskiy O, Watson LA, et al. (2017) Inhibition of p25/Cdk5 attenuates tauopathy in mouse and iPSC models of frontotemporal dementia. J Neurosci 37: 9917-9924. doi: 10.1523/JNEUROSCI.0621-17.2017
|
| [157] |
Morgan BP (2018) Complement in the pathogenesis of Alzheimer's disease. Semin Immunopathol 40: 113-124. doi: 10.1007/s00281-017-0662-9
|
| [158] |
Bonham LW, Desikan RS, Yokoyama JS, et al. (2016) The relationship between complement factor C3, APOE epsilon4, amyloid and tau in Alzheimer's disease. Acta Neuropathol Commun 4: 65. doi: 10.1186/s40478-016-0339-y
|
| [159] |
Bonham LW, Geier EG, Fan CC, et al. (2016) Age-dependent effects of APOE epsilon4 in preclinical Alzheimer's disease. Ann Clin Transl Neurol 3: 668-677. doi: 10.1002/acn3.333
|
| [160] |
Fritzinger DC, Benjamin DE (2016) The complement system in neuropathic and postoperative pain. Open Pain J 9: 26-37. doi: 10.2174/1876386301609010026
|
| [161] |
Sheedy FJ, Grebe A, Rayner KJ, et al. (2013) CD36 coordinates NLRP3 inflammasome activation by facilitating intracellular nucleation of soluble ligands into particulate ligands in sterile inflammation. Nat Immunol 14: 812-820. doi: 10.1038/ni.2639
|
| [162] |
Devanand DP (2018) Viral hypothesis and antiviral treatment in Alzheimer's disease. Curr Neurol Neurosci Rep 18: 55. doi: 10.1007/s11910-018-0863-1
|
| [163] | Duggan MR, Torkzaban B, Ahooyi TM, et al. (2020) Potential role for herpesviruses in Alzheimer's disease. J Alzheimers Dis 1-15. (Preprint). |
| [164] |
Wang JH, Cheng XR, Zhang XR, et al. (2016) Neuroendocrine immunomodulation network dysfunction in SAMP8 mice and PrP-hAbetaPPswe/PS1DeltaE9 mice: potential mechanism underlying cognitive impairment. Oncotarget 7: 22988-3005. doi: 10.18632/oncotarget.8453
|
| [165] |
Lopategui Cabezas I, Herrera Batista A, Penton Rol G (2014) The role of glial cells in Alzheimer disease: potential therapeutic implications. Neurologia 29: 305-309. doi: 10.1016/j.nrl.2012.10.006
|
| [166] |
Birch AM (2014) The contribution of astrocytes to Alzheimer's disease. Biochem Soc Trans 42: 1316-1320. doi: 10.1042/BST20140171
|
| [167] |
Takata K, Kitamura Y, Saeki M, et al. (2010) Galantamine-induced amyloid-β clearance mediated via stimulation of microglial nicotinic acetylcholine receptors. J Biol Chem 285: 40180-40191. doi: 10.1074/jbc.M110.142356
|
| [168] |
Koenigsknecht-Talboo J, Landreth GE (2005) Microglial phagocytosis induced by fibrillar beta-amyloid and IgGs are differentially regulated by proinflammatory cytokines. J Neurosci 25: 8240-8249. doi: 10.1523/JNEUROSCI.1808-05.2005
|
| [169] | Xie J, Wang H, Lin T, et al. (2017) Microglia-Synapse pathways: promising therapeutic strategy for Alzheimer's disease. Biomed Res Int 2017: 2986460. |
| [170] |
Qi Y, Klyubin I, Harney SC, et al. (2014) Longitudinal testing of hippocampal plasticity reveals the onset and maintenance of endogenous human Ass-induced synaptic dysfunction in individual freely behaving pre-plaque transgenic rats: rapid reversal by anti-Ass agents. Acta Neuropathol Commun 2: 175. doi: 10.1186/s40478-014-0175-x
|
| [171] |
Lo AC, Iscru E, Blum D, et al. (2013) Amyloid and tau neuropathology differentially affect prefrontal synaptic plasticity and cognitive performance in mouse models of Alzheimer's disease. J Alzheimers Dis 37: 109-125. doi: 10.3233/JAD-122296
|
| [172] |
Giulian D, Baker TJ (1986) Characterization of ameboid microglia isolated from developing mammalian brain. J Neurosci 6: 2163-2178. doi: 10.1523/JNEUROSCI.06-08-02163.1986
|
| [173] |
Kreutzberg GW (1996) Microglia: a sensor for pathological events in the CNS. Trends Neurosci 19: 312-318. doi: 10.1016/0166-2236(96)10049-7
|
| [174] |
Mecha M, Carrillo-Salinas FJ, Feliu A, et al. (2016) Microglia activation states and cannabinoid system: therapeutic implications. Pharmacol Ther 166: 40-55. doi: 10.1016/j.pharmthera.2016.06.011
|
| [175] |
Henkel JS, Beers DR, Zhao W, et al. (2009) Microglia in ALS: the good, the bad, and the resting. J Neuroimmune Pharmacol 4: 389-398. doi: 10.1007/s11481-009-9171-5
|
| [176] |
Stansley B, Post J, Hensley K (2012) A comparative review of cell culture systems for the study of microglial biology in Alzheimer's disease. J Neuroinflammation 9: 115. doi: 10.1186/1742-2094-9-115
|
| [177] |
Acosta C, Anderson HD, Anderson CM (2017) Astrocyte dysfunction in Alzheimer disease. J Neurosci Res 95: 2430-2447. doi: 10.1002/jnr.24075
|
| [178] |
Sofroniew MV (2009) Molecular dissection of reactive astrogliosis and glial scar formation. Trends Neurosci 32: 638-647. doi: 10.1016/j.tins.2009.08.002
|
| [179] |
Ricci G, Volpi L, Pasquali L, et al. (2009) Astrocyte-neuron interactions in neurological disorders. J Biol Phys 35: 317-336. doi: 10.1007/s10867-009-9157-9
|
| [180] |
Mattsson N, Tosun D, Insel PS, et al. (2014) Association of brain amyloid-beta with cerebral perfusion and structure in Alzheimer's disease and mild cognitive impairment. Brain 137: 1550-1561. doi: 10.1093/brain/awu043
|
| [181] |
Huang KL, Lin KJ, Ho MY, et al. (2012) Amyloid deposition after cerebral hypoperfusion: evidenced on [(18)F]AV-45 positron emission tomography. J Neurol Sci 319: 124-129. doi: 10.1016/j.jns.2012.04.014
|
| [182] |
Okamoto Y, Yamamoto T, Kalaria RN, et al. (2012) Cerebral hypoperfusion accelerates cerebral amyloid angiopathy and promotes cortical microinfarcts. Acta Neuropathol 123: 381-394. doi: 10.1007/s00401-011-0925-9
|
| [183] |
Pietronigro EC, Della Bianca V, Zenaro E, et al. (2017) NETosis in Alzheimer's disease. Front Immunol 8: 211. doi: 10.3389/fimmu.2017.00211
|
| [184] |
Mantovani A, Cassatella MA, Costantini C, et al. (2011) Neutrophils in the activation and regulation of innate and adaptive immunity. Nat Rev Immunol 11: 519-531. doi: 10.1038/nri3024
|
| [185] |
Tillack K, Breiden P, Martin R, et al. (2012) T lymphocyte priming by neutrophil extracellular traps links innate and adaptive immune responses. J Immunol 188: 3150-3159. doi: 10.4049/jimmunol.1103414
|
| [186] |
Llanos-Gonzalez E, Henares-Chavarino AA, Pedrero-Prieto CM, et al. (2019) Interplay between mitochondrial oxidative disorders and proteostasis in Alzheimer's disease. Front Neurosci 13: 1444. doi: 10.3389/fnins.2019.01444
|
| [187] |
Franco R, Vargas MR (2018) Redox biology in neurological function, dysfunction, and aging. Antioxid Redox Signal 28: 1583-1586. doi: 10.1089/ars.2018.7509
|
| [188] |
Miller VM, Lawrence DA, Mondal TK, et al. (2009) Reduced glutathione is highly expressed in white matter and neurons in the unperturbed mouse brain—implications for oxidative stress associated with neurodegeneration. Brain Res 1276: 22-30. doi: 10.1016/j.brainres.2009.04.029
|
| [189] |
Munoz P, Ardiles AO, Perez-Espinosa B, et al. (2020) Redox modifications in synaptic components as biomarkers of cognitive status, in brain aging and disease. Mech Ageing Dev 189: 111250. doi: 10.1016/j.mad.2020.111250
|
| [190] |
Kumar A, Yegla B, Foster TC (2018) Redox signaling in neurotransmission and cognition during aging. Antioxid Redox Signal 28: 1724-1745. doi: 10.1089/ars.2017.7111
|
| [191] |
Sbodio JI, Snyder SH, Paul BD (2019) Redox mechanisms in neurodegeneration: from disease outcomes to therapeutic opportunities. Antioxid Redox Signal 30: 1450-1499. doi: 10.1089/ars.2017.7321
|
| [192] |
Arias-Cavieres A, Adasme T, Sanchez G, et al. (2017) Aging impairs hippocampal-dependent recognition memory and LTP and prevents the associated RyR Up-regulation. Front Aging Neurosci 9: 111. doi: 10.3389/fnagi.2017.00111
|
| [193] |
Wilson C, Gonzalez-Billault C (2015) Regulation of cytoskeletal dynamics by redox signaling and oxidative stress: implications for neuronal development and trafficking. Front Cell Neurosci 9: 381. doi: 10.3389/fncel.2015.00381
|
| [194] |
Quintanilla RA, Orellana JA, von Bernhardi R (2012) Understanding risk factors for Alzheimer's disease: interplay of neuroinflammation, connexin-based communication and oxidative stress. Arch Med Res 43: 632-644. doi: 10.1016/j.arcmed.2012.10.016
|
| [195] |
Butterfield DA, Bader Lange ML, Sultana R (2010) Involvements of the lipid peroxidation product, HNE, in the pathogenesis and progression of Alzheimer's disease. Biochim Biophys Acta 1801: 924-929. doi: 10.1016/j.bbalip.2010.02.005
|
| [196] |
Chen YY, Yu XY, Chen L, et al. (2019) Redox signaling in aging kidney and opportunity for therapeutic intervention through natural products. Free Radic Biol Med 141: 141-149. doi: 10.1016/j.freeradbiomed.2019.06.012
|
| [197] |
Bruce-Keller AJ, Gupta S, Knight AG, et al. (2011) Cognitive impairment in humanized APPxPS1 mice is linked to Abeta(1-42) and NOX activation. Neurobiol Dis 44: 317-326. doi: 10.1016/j.nbd.2011.07.012
|
| [198] |
Kothari V, Luo Y, Tornabene T, et al. (2017) High fat diet induces brain insulin resistance and cognitive impairment in mice. Biochim Biophys Acta 1863: 499-508. doi: 10.1016/j.bbadis.2016.10.006
|
| [199] |
Wilkinson BL, Landreth GE (2006) The microglial NADPH oxidase complex as a source of oxidative stress in Alzheimer's disease. J Neuroinflammation 3: 30. doi: 10.1186/1742-2094-3-30
|
| [200] |
Wong KY, Roy J, Fung ML, et al. (2020) Relationships between mitochondrial dysfunction and neurotransmission failure in Alzheimer's disease. Aging Dis 11: 1291-1316. doi: 10.14336/AD.2019.1125
|
| [201] |
Bissette G, Seidler FJ, Nemeroff CB, et al. (1996) High affinity choline transporter status in Alzheimer's disease tissue from rapid autopsy. Ann N Y Acad Sci 777: 197-204. doi: 10.1111/j.1749-6632.1996.tb34419.x
|
| [202] |
Campanucci VA, Krishnaswamy A, Cooper E (2008) Mitochondrial reactive oxygen species inactivate neuronal nicotinic acetylcholine receptors and induce long-term depression of fast nicotinic synaptic transmission. J Neurosci 28: 1733-1744. doi: 10.1523/JNEUROSCI.5130-07.2008
|
| [203] |
Jomova K, Vondrakova D, Lawson M, et al. (2010) Metals, oxidative stress and neurodegenerative disorders. Mol Cell Biochem 345: 91-104. doi: 10.1007/s11010-010-0563-x
|
| [204] |
Swerdlow RH, Khan SM (2004) A “mitochondrial cascade hypothesis” for sporadic Alzheimer's disease. Med Hypotheses 63: 8-20. doi: 10.1016/j.mehy.2003.12.045
|
| [205] |
Kim GW, Gasche Y, Grzeschik S, et al. (2003) Neurodegeneration in striatum induced by the mitochondrial toxin 3-nitropropionic acid: role of matrix metalloproteinase-9 in early blood-brain barrier disruption? J Neurosci 23: 8733-8742. doi: 10.1523/JNEUROSCI.23-25-08733.2003
|
| [206] |
Ridnour LA, Dhanapal S, Hoos M, et al. (2012) Nitric oxide-mediated regulation of beta-amyloid clearance via alterations of MMP-9/TIMP-1. J Neurochem 123: 736-749. doi: 10.1111/jnc.12028
|
| [207] |
Liu Y, Liu F, Iqbal K, et al. (2008) Decreased glucose transporters correlate to abnormal hyperphosphorylation of tau in Alzheimer disease. FEBS Lett 582: 359-364. doi: 10.1016/j.febslet.2007.12.035
|
| [208] |
Nikinmaa M, Pursiheimo S, Soitamo AJ (2004) Redox state regulates HIF-1alpha and its DNA binding and phosphorylation in salmonid cells. J Cell Sci 117: 3201-3206. doi: 10.1242/jcs.01192
|
| [209] |
Morris G, Walder K, Puri BK, et al. (2016) The deleterious effects of oxidative and nitrosative stress on palmitoylation, membrane lipid rafts and lipid-based cellular signalling: new drug targets in neuroimmune disorders. Mol Neurobiol 53: 4638-4658. doi: 10.1007/s12035-015-9392-y
|
| [210] |
Zou Y, Watters A, Cheng N, et al. (2019) Polyunsaturated fatty acids from astrocytes activate PPARgamma signaling in cancer cells to promote brain metastasis. Cancer Discov 9: 1720-1735. doi: 10.1158/2159-8290.CD-19-0270
|
| [211] |
Marnett LJ (1999) Lipid peroxidation-DNA damage by malondialdehyde. Mutat Res 424: 83-95. doi: 10.1016/S0027-5107(99)00010-X
|
| [212] |
Lu Y, Ren J, Cui S, et al. (2016) Cerebral glucose metabolism assessment in rat models of Alzheimer's disease: an 18F-FDG-PET study. Am J Alzheimers Dis Other Demen 31: 333-340. doi: 10.1177/1533317515617725
|
| [213] |
Jeong DU, Oh JH, Lee JE, et al. (2016) Basal forebrain cholinergic deficits reduce glucose metabolism and function of cholinergic and GABAergic systems in the cingulate cortex. Yonsei Med J 57: 165-172. doi: 10.3349/ymj.2016.57.1.165
|
| [214] |
Braak H, Del Tredici K (2015) The preclinical phase of the pathological process underlying sporadic Alzheimer's disease. Brain 138: 2814-2833. doi: 10.1093/brain/awv236
|
| [215] |
Morrison BM, Lee Y, Rothstein JD (2013) Oligodendroglia: metabolic supporters of axons. Trends Cell Biol 23: 644-651. doi: 10.1016/j.tcb.2013.07.007
|
| [216] |
Grant CM, Quinn KA, Dawes IW (1999) Differential protein S-thiolation of glyceraldehyde-3-phosphate dehydrogenase isoenzymes influences sensitivity to oxidative stress. Mol Cell Biol 19: 2650-2656. doi: 10.1128/MCB.19.4.2650
|
| [217] |
Avery SV (2011) Molecular targets of oxidative stress. Biochem J 434: 201-210. doi: 10.1042/BJ20101695
|
| [218] |
Bubber P, Haroutunian V, Fisch G, et al. (2005) Mitochondrial abnormalities in Alzheimer brain: mechanistic implications. Ann Neurol 57: 695-703. doi: 10.1002/ana.20474
|
| [219] |
Gibson GE, Blass JP, Beal MF, et al. (2005) The alpha-ketoglutarate-dehydrogenase complex: a mediator between mitochondria and oxidative stress in neurodegeneration. Mol Neurobiol 31: 43-63. doi: 10.1385/MN:31:1-3:043
|
| [220] |
Long J, Liu C, Sun L, et al. (2009) Neuronal mitochondrial toxicity of malondialdehyde: inhibitory effects on respiratory function and enzyme activities in rat brain mitochondria. Neurochem Res 34: 786-794. doi: 10.1007/s11064-008-9882-7
|
| [221] |
Martin E, Rosenthal RE, Fiskum G (2005) Pyruvate dehydrogenase complex: metabolic link to ischemic brain injury and target of oxidative stress. J Neurosci Res 79: 240-247. doi: 10.1002/jnr.20293
|
| [222] |
Salminen A, Haapasalo A, Kauppinen A, et al. (2015) Impaired mitochondrial energy metabolism in Alzheimer's disease: impact on pathogenesis via disturbed epigenetic regulation of chromatin landscape. Prog Neurobiol 131: 1-20. doi: 10.1016/j.pneurobio.2015.05.001
|
| [223] |
Liguori C, Chiaravalloti A, Sancesario G, et al. (2016) Cerebrospinal fluid lactate levels and brain [18F]FDG PET hypometabolism within the default mode network in Alzheimer's disease. Eur J Nucl Med Mol Imaging 43: 2040-2049. doi: 10.1007/s00259-016-3417-2
|
| [224] |
Isopi E, Granzotto A, Corona C, et al. (2015) Pyruvate prevents the development of age-dependent cognitive deficits in a mouse model of Alzheimer's disease without reducing amyloid and tau pathology. Neurobiol Dis 81: 214-224. doi: 10.1016/j.nbd.2014.11.013
|
| [225] |
Talbot K, Wang HY, Kazi H, et al. (2012) Demonstrated brain insulin resistance in Alzheimer's disease patients is associated with IGF-1 resistance, IRS-1 dysregulation, and cognitive decline. J Clin Invest 122: 1316-1338. doi: 10.1172/JCI59903
|
| [226] |
Morales-Corraliza J, Wong H, Mazzella MJ, et al. (2016) Brain-Wide insulin resistance, Tau phosphorylation changes, and hippocampal neprilysin and Amyloid-beta alterations in a monkey model of Type 1 diabetes. J Neurosci 36: 4248-4258. doi: 10.1523/JNEUROSCI.4640-14.2016
|
| [227] |
Yamamoto N, Ishikuro R, Tanida M, et al. (2018) Insulin-signaling pathway regulates the degradation of amyloid beta-protein via astrocytes. Neuroscience 385: 227-236. doi: 10.1016/j.neuroscience.2018.06.018
|
| [228] |
Han X, Yang L, Du H, et al. (2016) Insulin attenuates beta-amyloid-associated Insulin/Akt/EAAT signaling perturbations in human astrocytes. Cell Mol Neurobiol 36: 851-864. doi: 10.1007/s10571-015-0268-5
|
| [229] |
Ng RC, Chan KH (2017) Potential neuroprotective effects of adiponectin in Alzheimer's disease. Int J Mol Sci 18: 592. doi: 10.3390/ijms18030592
|
| [230] |
Pei JJ, Khatoon S, An WL, et al. (2003) Role of protein kinase B in Alzheimer's neurofibrillary pathology. Acta Neuropathol 105: 381-392. doi: 10.1007/s00401-002-0657-y
|
| [231] |
Anderson NJ, King MR, Delbruck L, et al. (2014) Role of insulin signaling impairment, adiponectin and dyslipidemia in peripheral and central neuropathy in mice. Dis Model Mech 7: 625-633. doi: 10.1242/dmm.015750
|
| [232] |
Garcia-Casares N, Jorge RE, Garcia-Arnes JA, et al. (2014) Cognitive dysfunctions in middle-aged type 2 diabetic patients and neuroimaging correlations: a cross-sectional study. J Alzheimers Dis 42: 1337-1346. doi: 10.3233/JAD-140702
|
| [233] |
Purnell C, Gao S, Callahan CM, et al. (2009) Cardiovascular risk factors and incident Alzheimer disease: a systematic review of the literature. Alzheimer Dis Assoc Disord 23: 1-10. doi: 10.1097/WAD.0b013e318187541c
|
| [234] |
de la Torre JC (2012) Cardiovascular risk factors promote brain hypoperfusion leading to cognitive decline and dementia. Cardiovasc Psychiatry Neurol 2012: 367516. doi: 10.1155/2012/367516
|
| [235] |
Klohs J (2019) An integrated view on vascular dysfunction in Alzheimer's disease. Neurodegener Dis 19: 109-127. doi: 10.1159/000505625
|
| [236] |
Popovic M, Laumonnier Y, Burysek L, et al. (2008) Thrombin-induced expression of endothelial CX3CL1 potentiates monocyte CCL2 production and transendothelial migration. J Leukoc Biol 84: 215-223. doi: 10.1189/jlb.0907652
|
| [237] |
Sole M, Esteban-Lopez M, Taltavull B, et al. (2019) Blood-brain barrier dysfunction underlying Alzheimer's disease is induced by an SSAO/VAP-1-dependent cerebrovascular activation with enhanced Abeta deposition. Biochim Biophys Acta Mol Basis Dis 1865: 2189-2202. doi: 10.1016/j.bbadis.2019.04.016
|
| [238] |
Zlokovic BV (2011) Neurovascular pathways to neurodegeneration in Alzheimer's disease and other disorders. Nat Rev Neurosci 12: 723-738. doi: 10.1038/nrn3114
|
| [239] |
Ramanathan A, Nelson AR, Sagare AP, et al. (2015) Impaired vascular-mediated clearance of brain amyloid beta in Alzheimer's disease: the role, regulation and restoration of LRP1. Front Aging Neurosci 7: 136. doi: 10.3389/fnagi.2015.00136
|
| [240] |
Montagne A, Pa J, Zlokovic BV (2015) Vascular plasticity and cognition during normal aging and dementia. JAMA Neurol 72: 495-496. doi: 10.1001/jamaneurol.2014.4636
|
| [241] |
Montine TJ, Koroshetz WJ, Babcock D, et al. (2014) Recommendations of the Alzheimer's disease-related dementias conference. Neurology 83: 851-860. doi: 10.1212/WNL.0000000000000733
|
| [242] |
Sweeney MD, Sagare AP, Zlokovic BV (2015) Cerebrospinal fluid biomarkers of neurovascular dysfunction in mild dementia and Alzheimer's disease. J Cereb Blood Flow Metab 35: 1055-1068. doi: 10.1038/jcbfm.2015.76
|
| [243] |
Yan SD, Chen X, Fu J, et al. (1996) RAGE and amyloid-beta peptide neurotoxicity in Alzheimer's disease. Nature 382: 685-691. doi: 10.1038/382685a0
|
| [244] |
Miller MC, Tavares R, Johanson CE, et al. (2008) Hippocampal RAGE immunoreactivity in early and advanced Alzheimer's disease. Brain Res 1230: 273-280. doi: 10.1016/j.brainres.2008.06.124
|
| [245] |
Lue LF, Walker DG, Brachova L, et al. (2001) Involvement of microglial receptor for advanced glycation endproducts (RAGE) in Alzheimer's disease: identification of a cellular activation mechanism. Exp Neurol 171: 29-45. doi: 10.1006/exnr.2001.7732
|
| [246] |
Carnevale D, Mascio G, D'Andrea I, et al. (2012) Hypertension induces brain beta-amyloid accumulation, cognitive impairment, and memory deterioration through activation of receptor for advanced glycation end products in brain vasculature. Hypertension 60: 188-197. doi: 10.1161/HYPERTENSIONAHA.112.195511
|
| [247] |
Srikanth V, Maczurek A, Phan T, et al. (2011) Advanced glycation endproducts and their receptor RAGE in Alzheimer's disease. Neurobiol Aging 32: 763-777. doi: 10.1016/j.neurobiolaging.2009.04.016
|
| [248] |
de la Torre J (2018) The vascular hypothesis of Alzheimer's disease: a key to preclinical prediction of dementia using neuroimaging. J Alzheimers Dis 63: 35-52. doi: 10.3233/JAD-180004
|
| [249] |
Wierenga CE, Hays CC, Zlatar ZZ (2014) Cerebral blood flow measured by arterial spin labeling MRI as a preclinical marker of Alzheimer's disease. J Alzheimers Dis 42: S411-419. doi: 10.3233/JAD-141467
|
| [250] |
Chuang YF, Breitner JCS, Chiu YL, et al. (2014) Use of diuretics is associated with reduced risk of Alzheimer's disease: the cache county study. Neurobiol Aging 35: 2429-2435. doi: 10.1016/j.neurobiolaging.2014.05.002
|
| [251] |
Ashby EL, Kehoe PG (2013) Current status of renin-aldosterone angiotensin system-targeting anti-hypertensive drugs as therapeutic options for Alzheimer's disease. Expert Opin Investig Drugs 22: 1229-1242. doi: 10.1517/13543784.2013.812631
|
| [252] |
Yasar S, Xia J, Yao W, et al. (2013) Antihypertensive drugs decrease risk of Alzheimer disease: ginkgo evaluation of memory study. Neurology 81: 896-903. doi: 10.1212/WNL.0b013e3182a35228
|
| [253] |
Thomas T, Miners S, Love S (2015) Post-mortem assessment of hypoperfusion of cerebral cortex in Alzheimer's disease and vascular dementia. Brain 138: 1059-1069. doi: 10.1093/brain/awv025
|
| [254] |
Zhao Y, Gong CX (2015) From chronic cerebral hypoperfusion to Alzheimer-like brain pathology and neurodegeneration. Cell Mol Neurobiol 35: 101-110. doi: 10.1007/s10571-014-0127-9
|
| [255] |
Glodzik L, Rusinek H, Pirraglia E, et al. (2014) Blood pressure decrease correlates with tau pathology and memory decline in hypertensive elderly. Neurobiol Aging 35: 64-71. doi: 10.1016/j.neurobiolaging.2013.06.011
|
| [256] |
Hong S, Beja-Glasser VF, Nfonoyim BM, et al. (2016) Complement and microglia mediate early synapse loss in Alzheimer mouse models. Science 352: 712-716. doi: 10.1126/science.aad8373
|
| [257] |
Gottfries CG, Bartfai T, Carlsson A, et al. (1986) Multiple biochemical deficits in both gray and white matter of Alzheimer brains. Prog Neuropsychopharmacol Biol Psychiatry 10: 405-413. doi: 10.1016/0278-5846(86)90014-X
|
| [258] |
Storga D, Vrecko K, Birkmayer JG, et al. (1996) Monoaminergic neurotransmitters, their precursors and metabolites in brains of Alzheimer patients. Neurosci Lett 203: 29-32. doi: 10.1016/0304-3940(95)12256-7
|
| [259] |
Arai H, Ichimiya Y, Kosaka K, et al. (1992) Neurotransmitter changes in early- and late-onset Alzheimer-type dementia. Prog Neuropsychopharmacol Biol Psychiatry 16: 883-890. doi: 10.1016/0278-5846(92)90106-O
|
| [260] |
Lanari A, Amenta F, Silvestrelli G, et al. (2006) Neurotransmitter deficits in behavioural and psychological symptoms of Alzheimer's disease. Mech Ageing Dev 127: 158-165. doi: 10.1016/j.mad.2005.09.016
|
| [261] |
Nobili A, Latagliata EC, Viscomi MT, et al. (2017) Dopamine neuronal loss contributes to memory and reward dysfunction in a model of Alzheimer's disease. Nat Commun 8: 14727. doi: 10.1038/ncomms14727
|
| [262] |
Chalermpalanupap T, Kinkead B, Hu WT, et al. (2013) Targeting norepinephrine in mild cognitive impairment and Alzheimer's disease. Alzheimers Res Ther 5: 21. doi: 10.1186/alzrt175
|
| [263] |
Woolf NJ, Butcher LL (2011) Cholinergic systems mediate action from movement to higher consciousness. Behav Brain Res 221: 488-498. doi: 10.1016/j.bbr.2009.12.046
|
| [264] |
Baker-Nigh A, Vahedi S, Davis EG, et al. (2015) Neuronal amyloid-beta accumulation within cholinergic basal forebrain in ageing and Alzheimer's disease. Brain 138: 1722-1737. doi: 10.1093/brain/awv024
|
| [265] |
Bracco L, Bessi V, Padiglioni S, et al. (2014) Do cholinesterase inhibitors act primarily on attention deficit? A naturalistic study in Alzheimer's disease patients. J Alzheimers Dis 40: 737-742. doi: 10.3233/JAD-131154
|
| [266] |
Stephan AH, Barres BA, Stevens B (2012) The complement system: an unexpected role in synaptic pruning during development and disease. Annu Rev Neurosci 35: 369-389. doi: 10.1146/annurev-neuro-061010-113810
|
| [267] |
Bialas AR, Stevens B (2013) TGF-beta signaling regulates neuronal C1q expression and developmental synaptic refinement. Nat Neurosci 16: 1773-1782. doi: 10.1038/nn.3560
|
| [268] |
Nakajima K, Tohyama Y, Maeda S, et al. (2007) Neuronal regulation by which microglia enhance the production of neurotrophic factors for GABAergic, catecholaminergic, and cholinergic neurons. Neurochem Int 50: 807-820. doi: 10.1016/j.neuint.2007.02.006
|
| [269] |
Sierra A, Gottfried-Blackmore AC, McEwen BS, et al. (2007) Microglia derived from aging mice exhibit an altered inflammatory profile. Glia 55: 412-424. doi: 10.1002/glia.20468
|
| [270] |
Welser-Alves JV, Milner R (2013) Microglia are the major source of TNF-alpha and TGF-beta1 in postnatal glial cultures; regulation by cytokines, lipopolysaccharide, and vitronectin. Neurochem Int 63: 47-53. doi: 10.1016/j.neuint.2013.04.007
|
| [271] | Brucato FH, Benjamin DE (2020) Synaptic pruning in Alzheimer's disease: role of the complement system. Glob J Med Res 20. |
| [272] |
Gajardo I, Salazar CS, Lopez-Espindola D, et al. (2018) Lack of pannexin 1 alters synaptic GluN2 subunit composition and spatial reversal learning in mice. Front Mol Neurosci 11: 114. doi: 10.3389/fnmol.2018.00114
|
| [273] |
Flores-Munoz C, Gomez B, Mery E, et al. (2020) Acute pannexin 1 blockade mitigates early synaptic plasticity defects in a mouse model of Alzheimer's disease. Front Cell Neurosci 14: 46. doi: 10.3389/fncel.2020.00046
|
| [274] |
Liu J, Chang L, Song Y, et al. (2019) The role of NMDA receptors in Alzheimer's disease. Front Neurosci 13: 43. doi: 10.3389/fnins.2019.00043
|
| [275] |
Hidalgo C, Arias-Cavieres A (2016) Calcium, reactive oxygen species, and synaptic plasticity. Physiology 31: 201-215. doi: 10.1152/physiol.00038.2015
|
| [276] |
Lu YF, Hawkins RD (2002) Ryanodine receptors contribute to cGMP-induced late-phase LTP and CREB phosphorylation in the hippocampus. J Neurophysiol 88: 1270-1278. doi: 10.1152/jn.2002.88.3.1270
|
| [277] |
Del Prete D, Checler F, Chami M (2014) Ryanodine receptors: physiological function and deregulation in Alzheimer disease. Mol Neurodegener 9: 21. doi: 10.1186/1750-1326-9-21
|
| [278] |
Oules B, Del Prete D, Greco B, et al. (2012) Ryanodine receptor blockade reduces amyloid-beta load and memory impairments in Tg2576 mouse model of Alzheimer disease. J Neurosci 32: 11820-11834. doi: 10.1523/JNEUROSCI.0875-12.2012
|
| [279] |
SanMartin CD, Veloso P, Adasme T, et al. (2017) RyR2-Mediated Ca(2+) release and mitochondrial ros generation partake in the synaptic dysfunction caused by amyloid beta peptide oligomers. Front Mol Neurosci 10: 115. doi: 10.3389/fnmol.2017.00115
|
| [280] |
Munoz Y, Paula-Lima AC, Nunez MT (2018) Reactive oxygen species released from astrocytes treated with amyloid beta oligomers elicit neuronal calcium signals that decrease phospho-Ser727-STAT3 nuclear content. Free Radic Biol Med 117: 132-144. doi: 10.1016/j.freeradbiomed.2018.01.006
|
| [281] |
Bitzer-Quintero OK, Gonzalez-Burgos I (2012) Immune system in the brain: a modulatory role on dendritic spine morphophysiology? Neural Plast 2012: 348642. doi: 10.1155/2012/348642
|
| [282] |
Fonseca MI, Chu SH, Hernandez MX, et al. (2017) Cell-specific deletion of C1qa identifies microglia as the dominant source of C1q in mouse brain. J Neuroinflammation 14: 48. doi: 10.1186/s12974-017-0814-9
|
| [283] |
Schafer DP, Lehrman EK, Kautzman AG, et al. (2012) Microglia sculpt postnatal neural circuits in an activity and complement-dependent manner. Neuron 74: 691-705. doi: 10.1016/j.neuron.2012.03.026
|
| [284] |
Gasque P (2004) Complement: a unique innate immune sensor for danger signals. Mol Immunol 41: 1089-1098. doi: 10.1016/j.molimm.2004.06.011
|
| [285] |
Azevedo EP, Ledo JH, Barbosa G, et al. (2013) Activated microglia mediate synapse loss and short-term memory deficits in a mouse model of transthyretin-related oculoleptomeningeal amyloidosis. Cell Death Dis 4: e789. doi: 10.1038/cddis.2013.325
|
| [286] | Wang WY, Tan MS, Yu JT, et al. (2015) Role of pro-inflammatory cytokines released from microglia in Alzheimer's disease. Ann Transl Med 3: 136. |
| [287] |
Liddelow SA, Guttenplan KA, Clarke LE, et al. (2017) Neurotoxic reactive astrocytes are induced by activated microglia. Nature 541: 481-487. doi: 10.1038/nature21029
|
| [288] |
Stevens B, Allen NJ, Vazquez LE, et al. (2007) The classical complement cascade mediates CNS synapse elimination. Cell 131: 1164-1178. doi: 10.1016/j.cell.2007.10.036
|
| [289] |
Stephan AH, Madison DV, Mateos JM, et al. (2013) A dramatic increase of C1q protein in the CNS during normal aging. J Neurosci 33: 13460-13474. doi: 10.1523/JNEUROSCI.1333-13.2013
|
| [290] |
Shi Q, Colodner KJ, Matousek SB, et al. (2015) Complement C3-Deficient mice fail to display age-related hippocampal decline. J Neurosci 35: 13029-13042. doi: 10.1523/JNEUROSCI.1698-15.2015
|
| [291] |
Shi Q, Chowdhury S, Ma R, et al. (2017) Complement C3 deficiency protects against neurodegeneration in aged plaque-rich APP/PS1 mice. Sci Transl Med 9: eaaf6295. doi: 10.1126/scitranslmed.aaf6295
|
| [292] |
Reichwald J, Danner S, Wiederhold KH, et al. (2009) Expression of complement system components during aging and amyloid deposition in APP transgenic mice. J Neuroinflammation 6: 35. doi: 10.1186/1742-2094-6-35
|
| [293] |
Paolicelli RC, Jawaid A, Henstridge CM, et al. (2017) TDP-43 depletion in microglia promotes amyloid clearance but also induces synapse loss. Neuron 95: 297-308. e6. doi: 10.1016/j.neuron.2017.05.037
|
| [294] |
Orellana JA, Shoji KF, Abudara V, et al. (2011) Amyloid beta-induced death in neurons involves glial and neuronal hemichannels. J Neurosci 31: 4962-4977. doi: 10.1523/JNEUROSCI.6417-10.2011
|
| [295] |
Pena-Oyarzun D, Bravo-Sagua R, Diaz-Vega A, et al. (2018) Autophagy and oxidative stress in non-communicable diseases: a matter of the inflammatory state? Free Radic Biol Med 124: 61-78. doi: 10.1016/j.freeradbiomed.2018.05.084
|
| [296] | Mizushima N (2009) Physiological functions of autophagy. Curr Top Microbiol Immunol 335: 71-84. |
| [297] |
Alirezaei M, Kiosses WB, Flynn CT, et al. (2008) Disruption of neuronal autophagy by infected microglia results in neurodegeneration. PLoS One 3: e2906. doi: 10.1371/journal.pone.0002906
|
| [298] |
Komatsu M, Waguri S, Chiba T, et al. (2006) Loss of autophagy in the central nervous system causes neurodegeneration in mice. Nature 441: 880-884. doi: 10.1038/nature04723
|
| [299] |
Ji ZS, Mullendorff K, Cheng IH, et al. (2006) Reactivity of apolipoprotein E4 and amyloid beta peptide: lysosomal stability and neurodegeneration. J Biol Chem 281: 2683-2692. doi: 10.1074/jbc.M506646200
|
| [300] |
Belinson H, Lev D, Masliah E, et al. (2008) Activation of the amyloid cascade in apolipoprotein E4 transgenic mice induces lysosomal activation and neurodegeneration resulting in marked cognitive deficits. J Neurosci 28: 4690-4701. doi: 10.1523/JNEUROSCI.5633-07.2008
|
| [301] |
Friedman LG, Qureshi YH, Yu WH (2015) Promoting autophagic clearance: viable therapeutic targets in Alzheimer's disease. Neurotherapeutics 12: 94-108. doi: 10.1007/s13311-014-0320-z
|
| [302] |
Yu WH, Cuervo AM, Kumar A, et al. (2005) Macroautophagy—a novel Beta-amyloid peptide-generating pathway activated in Alzheimer's disease. J Cell Biol 171: 87-98. doi: 10.1083/jcb.200505082
|
| [303] |
Nixon RA (2007) Autophagy, amyloidogenesis and Alzheimer disease. J Cell Sci 120: 4081-4091. doi: 10.1242/jcs.019265
|
| [304] |
Klionsky DJ, Elazar Z, Seglen PO, et al. (2008) Does bafilomycin A1 block the fusion of autophagosomes with lysosomes? Autophagy 4: 849-850. doi: 10.4161/auto.6845
|
| [305] |
Nixon RA, Wegiel J, Kumar A, et al. (2005) Extensive involvement of autophagy in Alzheimer disease: an immuno-electron microscopy study. J Neuropathol Exp Neurol 64: 113-122. doi: 10.1093/jnen/64.2.113
|
| [306] |
Funderburk SF, Marcellino BK, Yue Z (2010) Cell “self-eating” (autophagy) mechanism in Alzheimer's disease. Mt Sinai J Med 77: 59-68. doi: 10.1002/msj.20161
|
| [307] |
Silva DF, Esteves AR, Oliveira CR, et al. (2011) Mitochondria: the common upstream driver of amyloid-beta and tau pathology in Alzheimer's disease. Curr Alzheimer Res 8: 563-572. doi: 10.2174/156720511796391872
|
| [308] |
Yu WH, Kumar A, Peterhoff C, et al. (2004) Autophagic vacuoles are enriched in amyloid precursor protein-secretase activities: implications for beta-amyloid peptide over-production and localization in Alzheimer's disease. Int J Biochem Cell Biol 36: 2531-2540. doi: 10.1016/j.biocel.2004.05.010
|
| [309] |
Mizushima N (2005) A(beta) generation in autophagic vacuoles. J Cell Biol 171: 15-17. doi: 10.1083/jcb.200508097
|
| [310] |
Boland B, Kumar A, Lee S, et al. (2008) Autophagy induction and autophagosome clearance in neurons: relationship to autophagic pathology in Alzheimer's disease. J Neurosci 28: 6926-6937. doi: 10.1523/JNEUROSCI.0800-08.2008
|
| [311] |
Lautrup S, Lou G, Aman Y, et al. (2019) Microglial mitophagy mitigates neuroinflammation in Alzheimer's disease. Neurochem Int 129: 104469. doi: 10.1016/j.neuint.2019.104469
|
| [312] |
Reddy PH, Oliver DM (2019) Amyloid Beta and phosphorylated Tau-Induced defective autophagy and mitophagy in Alzheimer's disease. Cells 8: 488. doi: 10.3390/cells8050488
|
| [313] |
Kerr JS, Adriaanse BA, Greig NH, et al. (2017) Mitophagy and Alzheimer's disease: cellular and molecular mechanisms. Trends Neurosci 40: 151-166. doi: 10.1016/j.tins.2017.01.002
|
| [314] |
Hu Y, Li XC, Wang ZH, et al. (2016) Tau accumulation impairs mitophagy via increasing mitochondrial membrane potential and reducing mitochondrial Parkin. Oncotarget 7: 17356-17368. doi: 10.18632/oncotarget.7861
|
| [315] |
Ellisdon AM, Bottomley SP (2004) The role of protein misfolding in the pathogenesis of human diseases. IUBMB Life 56: 119-123. doi: 10.1080/15216540410001674003
|
| [316] |
Vingtdeux V, Sergeant N, Buee L (2012) Potential contribution of exosomes to the prion-like propagation of lesions in Alzheimer's disease. Front Physiol 3: 229. doi: 10.3389/fphys.2012.00229
|
| [317] |
Uddin MS, Tewari D, Sharma G, et al. (2020) Molecular mechanisms of ER stress and UPR in the pathogenesis of Alzheimer's disease. Mol Neurobiol 57: 2902-2919. doi: 10.1007/s12035-020-01929-y
|
| [318] |
Schiera G, Di Liegro CM, Di Liegro I (2015) Extracellular membrane vesicles as vehicles for brain cell-to-cell interactions in physiological as well as pathological conditions. Biomed Res Int 2015: 152926. doi: 10.1155/2015/152926
|
| [319] |
Christianson JC, Ye Y (2014) Cleaning up in the endoplasmic reticulum: ubiquitin in charge. Nat Struct Mol Biol 21: 325-335. doi: 10.1038/nsmb.2793
|
| [320] |
Diehl JA, Fuchs SY, Koumenis C (2011) The cell biology of the unfolded protein response. Gastroenterology 141: 38-41. e2. doi: 10.1053/j.gastro.2011.05.018
|
| [321] |
Dhakal S, Macreadie I (2020) Protein homeostasis networks and the use of yeast to guide interventions in Alzheimer's disease. Int J Mol Sci 21: 8014. doi: 10.3390/ijms21218014
|
| [322] | Kabir MT, Uddin MS, Zaman S, et al. (2020) Molecular mechanisms of metal toxicity in the pathogenesis of Alzheimer's disease. Mol Neurobiol 1-20. |
| [323] |
Cristovao JS, Santos R, Gomes CM (2016) Metals and neuronal metal binding proteins implicated in Alzheimer's disease. Oxid Med Cell Longev 2016: 9812178. doi: 10.1155/2016/9812178
|
| [324] |
Gerber H, Wu F, Dimitrov M, et al. (2017) Zinc and copper differentially modulate amyloid precursor protein processing by gamma-secretase and amyloid-beta peptide production. J Biol Chem 292: 3751-3767. doi: 10.1074/jbc.M116.754101
|
| [325] |
Savelieff MG, Lee S, Liu Y, et al. (2013) Untangling amyloid-beta, tau, and metals in Alzheimer's disease. ACS Chem Biol 8: 856-865. doi: 10.1021/cb400080f
|
| [326] |
Dahms SO, Konnig I, Roeser D, et al. (2012) Metal binding dictates conformation and function of the amyloid precursor protein (APP) E2 domain. J Mol Biol 416: 438-452. doi: 10.1016/j.jmb.2011.12.057
|
| [327] |
Flaten TP (2001) Aluminium as a risk factor in Alzheimer's disease, with emphasis on drinking water. Brain Res Bull 55: 187-196. doi: 10.1016/S0361-9230(01)00459-2
|
| [328] |
Ward RJ, Zucca FA, Duyn JH, et al. (2014) The role of iron in brain ageing and neurodegenerative disorders. Lancet Neurol 13: 1045-1060. doi: 10.1016/S1474-4422(14)70117-6
|
| [329] |
Lane DJR, Ayton S, Bush AI (2018) Iron and Alzheimer's disease: an update on emerging mechanisms. J Alzheimers Dis 64: S379-S395. doi: 10.3233/JAD-179944
|
| [330] |
Tamano H, Takeda A (2011) Dynamic action of neurometals at the synapse. Metallomics 3: 656-661. doi: 10.1039/c1mt00008j
|
| [331] |
Ashraf A, Clark M, So PW (2018) The aging of iron man. Front Aging Neurosci 10: 65. doi: 10.3389/fnagi.2018.00065
|
| [332] |
Ojala JO, Sutinen EM (2017) The role of interleukin-18, oxidative stress and metabolic syndrome in Alzheimer's disease. J Clin Med 6: 55. doi: 10.3390/jcm6050055
|
| [333] |
Arrigoni F, Rizza F, Tisi R, et al. (2020) On the propagation of the OH radical produced by Cu-amyloid beta peptide model complexes. Insight from molecular modelling. Metallomics 12: 1765-1780. doi: 10.1039/D0MT00113A
|
| [334] |
Colvin RA, Jin Q, Lai B, et al. (2016) Visualizing metal content and intracellular distribution in primary hippocampal neurons with synchrotron X-ray fluorescence. PLoS One 11: e0159582. doi: 10.1371/journal.pone.0159582
|
| [335] |
Acevedo KM, Hung YH, Dalziel AH, et al. (2011) Copper promotes the trafficking of the amyloid precursor protein. J Biol Chem 286: 8252-8262. doi: 10.1074/jbc.M110.128512
|
| [336] |
Hickey JL, James JL, Henderson CA, et al. (2015) Intracellular distribution of fluorescent copper and zinc bis(thiosemicarbazonato) complexes measured with fluorescence lifetime spectroscopy. Inorg Chem 54: 9556-9567. doi: 10.1021/acs.inorgchem.5b01599
|
| [337] |
Alaverdashvili M, Hackett MJ, Pickering IJ, et al. (2014) Laminar-specific distribution of zinc: evidence for presence of layer IV in forelimb motor cortex in the rat. Neuroimage 103: 502-510. doi: 10.1016/j.neuroimage.2014.08.046
|
| [338] |
Ciccotosto GD, James SA, Altissimo M, et al. (2014) Quantitation and localization of intracellular redox active metals by X-ray fluorescence microscopy in cortical neurons derived from APP and APLP2 knockout tissue. Metallomics 6: 1894-1904. doi: 10.1039/C4MT00176A
|
| [339] |
Craddock TJ, Tuszynski JA, Chopra D, et al. (2012) The zinc dyshomeostasis hypothesis of Alzheimer's disease. PLoS One 7: e33552. doi: 10.1371/journal.pone.0033552
|
| [340] |
Christensen MK, Geneser FA (1995) Distribution of neurons of origin of zinc-containing projections in the amygdala of the rat. Anat Embryol 191: 227-237. doi: 10.1007/BF00187821
|
| [341] |
Jiang L, Dong H, Cao H, et al. (2019) Exosomes in pathogenesis, diagnosis, and treatment of Alzheimer's disease. Med Sci Monit 25: 3329-3335. doi: 10.12659/MSM.914027
|
| [342] |
Song Z, Xu Y, Deng W, et al. (2020) Brain derived exosomes are a double-edged sword in Alzheimer's disease. Front Mol Neurosci 13: 79. doi: 10.3389/fnmol.2020.00079
|
| [343] |
van Niel G, Porto-Carreiro I, Simoes S, et al. (2006) Exosomes: a common pathway for a specialized function. J Biochem 140: 13-21. doi: 10.1093/jb/mvj128
|
| [344] |
Eitan E, Suire C, Zhang S, et al. (2016) Impact of lysosome status on extracellular vesicle content and release. Ageing Res Rev 32: 65-74. doi: 10.1016/j.arr.2016.05.001
|
| [345] |
Asai H, Ikezu S, Tsunoda S, et al. (2015) Depletion of microglia and inhibition of exosome synthesis halt tau propagation. Nat Neurosci 18: 1584-1593. doi: 10.1038/nn.4132
|
| [346] |
Dinkins MB, Enasko J, Hernandez C, et al. (2016) Neutral sphingomyelinase-2 deficiency ameliorates Alzheimer's disease pathology and improves cognition in the 5XFAD mouse. J Neurosci 36: 8653-8667. doi: 10.1523/JNEUROSCI.1429-16.2016
|
| [347] |
Xiao T, Zhang W, Jiao B, et al. (2017) The role of exosomes in the pathogenesis of Alzheimer' disease. Transl Neurodegener 6: 3. doi: 10.1186/s40035-017-0072-x
|
| [348] |
Yuyama K, Igarashi Y (2017) Exosomes as carriers of Alzheimer's amyloid-ss. Front Neurosci 11: 229. doi: 10.3389/fnins.2017.00229
|
| [349] |
Cataldo AM, Barnett JL, Pieroni C, et al. (1997) Increased neuronal endocytosis and protease delivery to early endosomes in sporadic Alzheimer's disease: neuropathologic evidence for a mechanism of increased beta-amyloidogenesis. J Neurosci 17: 6142-6151. doi: 10.1523/JNEUROSCI.17-16-06142.1997
|
| [350] |
Cataldo A, Rebeck GW, Ghetri B, et al. (2001) Endocytic disturbances distinguish among subtypes of Alzheimer's disease and related disorders. Ann Neurol 50: 661-665. doi: 10.1002/ana.1254
|
| [351] |
Cataldo AM, Petanceska S, Terio NB, et al. (2004) Abeta localization in abnormal endosomes: association with earliest Abeta elevations in AD and Down syndrome. Neurobiol Aging 25: 1263-1272. doi: 10.1016/j.neurobiolaging.2004.02.027
|
| [352] |
Takahashi RH, Milner TA, Li F, et al. (2002) Intraneuronal Alzheimer abeta42 accumulates in multivesicular bodies and is associated with synaptic pathology. Am J Pathol 161: 1869-1879. doi: 10.1016/S0002-9440(10)64463-X
|
| [353] |
Langui D, Girardot N, El Hachimi KH, et al. (2004) Subcellular topography of neuronal Abeta peptide in APPxPS1 transgenic mice. Am J Pathol 165: 1465-1477. doi: 10.1016/S0002-9440(10)63405-0
|
| [354] |
Fiandaca MS, Kapogiannis D, Mapstone M, et al. (2015) Identification of preclinical Alzheimer's disease by a profile of pathogenic proteins in neurally derived blood exosomes: a case-control study. Alzheimers Dement 11: 600-607. e1. doi: 10.1016/j.jalz.2014.06.008
|
| [355] |
Wang G, Dinkins M, He Q, et al. (2012) Astrocytes secrete exosomes enriched with proapoptotic ceramide and prostate apoptosis response 4 (PAR-4): potential mechanism of apoptosis induction in Alzheimer disease (AD). J Biol Chem 287: 21384-21395. doi: 10.1074/jbc.M112.340513
|
| [356] |
Bindea G, Mlecnik B, Hackl H, et al. (2009) ClueGO: a Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 25: 1091-1093. doi: 10.1093/bioinformatics/btp101
|
| [357] |
Mosconi L, Berti V, Swerdlow RH, et al. (2010) Maternal transmission of Alzheimer's disease: prodromal metabolic phenotype and the search for genes. Hum Genomics 4: 170-193. doi: 10.1186/1479-7364-4-3-170
|
| [358] |
Birky CW (1995) Uniparental inheritance of mitochondrial and chloroplast genes: mechanisms and evolution. Proc Natl Acad Sci U S A 92: 11331-11338. doi: 10.1073/pnas.92.25.11331
|
| [359] |
Luo SM, Ge ZJ, Wang ZW, et al. (2013) Unique insights into maternal mitochondrial inheritance in mice. Proc Natl Acad Sci U S A 110: 13038-13043. doi: 10.1073/pnas.1303231110
|
| [360] |
Yu Z, O'Farrell PH, Yakubovich N, et al. (2017) The mitochondrial DNA polymerase promotes elimination of paternal mitochondrial genomes. Curr Biol 27: 1033-1039. doi: 10.1016/j.cub.2017.02.014
|
| [361] |
Boumezbeur F, Mason GF, de Graaf RA, et al. (2010) Altered brain mitochondrial metabolism in healthy aging as assessed by in vivo magnetic resonance spectroscopy. J Cereb Blood Flow Metab 30: 211-221. doi: 10.1038/jcbfm.2009.197
|
| [362] |
Rhein V, Song X, Wiesner A, et al. (2009) Amyloid-beta and tau synergistically impair the oxidative phosphorylation system in triple transgenic Alzheimer's disease mice. Proc Natl Acad Sci U S A 106: 20057-20062. doi: 10.1073/pnas.0905529106
|
| [363] |
Larosa V, Remacle C (2018) Insights into the respiratory chain and oxidative stress. Biosci Rep 38. doi: 10.1042/BSR20171492
|
| [364] |
Carden T, Singh B, Mooga V, et al. (2017) Epigenetic modification of miR-663 controls mitochondria-to-nucleus retrograde signaling and tumor progression. J Biol Chem 292: 20694-20706. doi: 10.1074/jbc.M117.797001
|
| [365] |
Spuch C, Ortolano S, Navarro C (2012) New insights in the amyloid-Beta interaction with mitochondria. J Aging Res 2012: 324968. doi: 10.1155/2012/324968
|
| [366] |
Yan SD, Stern DM (2005) Mitochondrial dysfunction and Alzheimer's disease: role of amyloid-beta peptide alcohol dehydrogenase (ABAD). Int J Exp Pathol 86: 161-171. doi: 10.1111/j.0959-9673.2005.00427.x
|
| [367] |
Cho DH, Nakamura T, Fang J, et al. (2009) S-nitrosylation of Drp1 mediates beta-amyloid-related mitochondrial fission and neuronal injury. Science 324: 102-105. doi: 10.1126/science.1171091
|
| [368] |
Area-Gomez E, Schon EA (2017) On the pathogenesis of Alzheimer's disease: the MAM Hypothesis. FASEB J 31: 864-867. doi: 10.1096/fj.201601309
|
| [369] |
Snowden SG, Ebshiana AA, Hye A, et al. (2017) Association between fatty acid metabolism in the brain and Alzheimer disease neuropathology and cognitive performance: a nontargeted metabolomic study. PLoS Med 14: e1002266. doi: 10.1371/journal.pmed.1002266
|
| [370] |
Kao YC, Ho PC, Tu YK, et al. (2020) Lipids and Alzheimer's disease. Int J Mol Sci 21: 1505. doi: 10.3390/ijms21041505
|
| [371] |
Czubowicz K, Jesko H, Wencel P, et al. (2019) The role of ceramide and Sphingosine-1-Phosphate in Alzheimer's disease and other neurodegenerative disorders. Mol Neurobiol 56: 5436-5455. doi: 10.1007/s12035-018-1448-3
|
| [372] |
Popugaeva E, Pchitskaya E, Bezprozvanny I (2018) Dysregulation of intracellular calcium signaling in Alzheimer's disease. Antioxid Redox Signal 29: 1176-1188. doi: 10.1089/ars.2018.7506
|
| [373] |
Ruiz A, Matute C, Alberdi E (2009) Endoplasmic reticulum Ca(2+) release through ryanodine and IP(3) receptors contributes to neuronal excitotoxicity. Cell Calcium 46: 273-281. doi: 10.1016/j.ceca.2009.08.005
|
| [374] |
Bezprozvanny I (2009) Calcium signaling and neurodegenerative diseases. Trends Mol Med 15: 89-100. doi: 10.1016/j.molmed.2009.01.001
|
| [375] |
Tong BC, Wu AJ, Li M, et al. (2018) Calcium signaling in Alzheimer's disease & therapies. Biochim Biophys Acta Mol Cell Res 1865: 1745-1760. doi: 10.1016/j.bbamcr.2018.07.018
|
| [376] |
Etcheberrigaray R, Hirashima N, Nee L, et al. (1998) Calcium responses in fibroblasts from asymptomatic members of Alzheimer's disease families. Neurobiol Dis 5: 37-45. doi: 10.1006/nbdi.1998.0176
|
| [377] |
Berridge MJ (2009) Inositol trisphosphate and calcium signalling mechanisms. Biochim Biophys Acta 1793: 933-940. doi: 10.1016/j.bbamcr.2008.10.005
|
| [378] |
Huang WJ, Zhang X, Chen WW (2016) Role of oxidative stress in Alzheimer's disease. Biomed Rep 4: 519-522. doi: 10.3892/br.2016.630
|
| [379] |
Mauvezin C, Neufeld TP (2015) Bafilomycin A1 disrupts autophagic flux by inhibiting both V-ATPase-dependent acidification and Ca-P60A/SERCA-dependent autophagosome-lysosome fusion. Autophagy 11: 1437-1438. doi: 10.1080/15548627.2015.1066957
|
| [380] |
Medina DL, Di Paola S, Peluso I, et al. (2015) Lysosomal calcium signalling regulates autophagy through calcineurin and TFEB. Nat Cell Biol 17: 288-299. doi: 10.1038/ncb3114
|
| [381] |
Mondal AC (2019) Role of GPCR signaling and calcium dysregulation in Alzheimer's disease. Mol Cell Neurosci 101: 103414. doi: 10.1016/j.mcn.2019.103414
|
| [382] |
Fernandez-Fernandez D, Rosenbrock H, Kroker KS (2015) Inhibition of PDE2A, but not PDE9A, modulates presynaptic short-term plasticity measured by paired-pulse facilitation in the CA1 region of the hippocampus. Synapse 69: 484-496. doi: 10.1002/syn.21840
|
| [383] | Zhang G, Stackman RW (2015) The role of serotonin 5-HT2A receptors in memory and cognition. Front Pharmacol 6: 225. |
| [384] | Raote I, Bhattacharya A, Panicker MM (2007) Serotonin 2A (5-HT2A) receptor function: ligand-dependent mechanisms and pathways. Serotonin Receptors in Neurobiology Boca Raton (FL): (Frontiers in Neuroscience), 105-132. |
| [385] |
Chang L, Karin M (2001) Mammalian MAP kinase signalling cascades. Nature 410: 37-40. doi: 10.1038/35065000
|
| [386] |
Hullinger R, O'Riordan K, Burger C (2015) Environmental enrichment improves learning and memory and long-term potentiation in young adult rats through a mechanism requiring mGluR5 signaling and sustained activation of p70s6k. Neurobiol Learn Mem 125: 126-134. doi: 10.1016/j.nlm.2015.08.006
|
| [387] |
Allen KD, Gourov AV, Harte C, et al. (2014) Nucleolar integrity is required for the maintenance of long-term synaptic plasticity. PLoS One 9: e104364. doi: 10.1371/journal.pone.0104364
|
| [388] |
Borroto-Escuela DO, Tarakanov AO, Guidolin D, et al. (2011) Moonlighting characteristics of G protein-coupled receptors: focus on receptor heteromers and relevance for neurodegeneration. IUBMB Life 63: 463-472. doi: 10.1002/iub.473
|
| [389] |
Spilman P, Podlutskaya N, Hart MJ, et al. (2010) Inhibition of mTOR by rapamycin abolishes cognitive deficits and reduces amyloid-beta levels in a mouse model of Alzheimer's disease. PLoS One 5: e9979. doi: 10.1371/journal.pone.0009979
|
| [390] |
Caccamo A, Maldonado MA, Majumder S, et al. (2011) Naturally secreted amyloid-beta increases mammalian target of rapamycin (mTOR) activity via a PRAS40-mediated mechanism. J Biol Chem 286: 8924-8932. doi: 10.1074/jbc.M110.180638
|
| [391] |
Lipton JO, Sahin M (2014) The neurology of mTOR. Neuron 84: 275-291. doi: 10.1016/j.neuron.2014.09.034
|
| [392] |
Oddo S (2012) The role of mTOR signaling in Alzheimer disease. Front Biosci 4: 941-952. doi: 10.2741/s310
|
| [393] |
Caccamo A, De Pinto V, Messina A, et al. (2014) Genetic reduction of mammalian target of rapamycin ameliorates Alzheimer's disease-like cognitive and pathological deficits by restoring hippocampal gene expression signature. J Neurosci 34: 7988-7998. doi: 10.1523/JNEUROSCI.0777-14.2014
|
| [394] |
Hodges SL, Reynolds CD, Smith GD, et al. (2018) Molecular interplay between hyperactive mammalian target of rapamycin signaling and Alzheimer's disease neuropathology in the NS-Pten knockout mouse model. Neuroreport 29: 1109-1113. doi: 10.1097/WNR.0000000000001081
|
| [395] |
Gabbouj S, Ryhanen S, Marttinen M, et al. (2019) Altered insulin signaling in Alzheimer's disease brain—special emphasis on PI3K-Akt Pathway. Front Neurosci 13: 629. doi: 10.3389/fnins.2019.00629
|
| [396] |
Magri L, Cambiaghi M, Cominelli M, et al. (2011) Sustained activation of mTOR pathway in embryonic neural stem cells leads to development of tuberous sclerosis complex-associated lesions. Cell Stem Cell 9: 447-462. doi: 10.1016/j.stem.2011.09.008
|
| [397] |
Li YH, Werner H, Puschel AW (2008) Rheb and mTOR regulate neuronal polarity through Rap1B. J Biol Chem 283: 33784-33792. doi: 10.1074/jbc.M802431200
|
| [398] |
Urbanska M, Gozdz A, Swiech LJ, et al. (2012) Mammalian target of rapamycin complex 1 (mTORC1) and 2 (mTORC2) control the dendritic arbor morphology of hippocampal neurons. J Biol Chem 287: 30240-30256. doi: 10.1074/jbc.M112.374405
|
| [399] |
Franco R, Martinez-Pinilla E, Navarro G, et al. (2017) Potential of GPCRs to modulate MAPK and mTOR pathways in Alzheimer's disease. Prog Neurobiol 149: 21-38. doi: 10.1016/j.pneurobio.2017.01.004
|
| [400] |
Perluigi M, Di Domenico F, Butterfield DA (2015) mTOR signaling in aging and neurodegeneration: at the crossroad between metabolism dysfunction and impairment of autophagy. Neurobiol Dis 84: 39-49. doi: 10.1016/j.nbd.2015.03.014
|
| [401] |
Ma T, Hoeffer CA, Capetillo-Zarate E, et al. (2010) Dysregulation of the mTOR pathway mediates impairment of synaptic plasticity in a mouse model of Alzheimer's disease. PLoS One 5: e12845. doi: 10.1371/journal.pone.0012845
|
| [402] |
Kudo W, Lee HP, Smith MA, et al. (2012) Inhibition of Bax protects neuronal cells from oligomeric Abeta neurotoxicity. Cell Death Dis 3: e309. doi: 10.1038/cddis.2012.43
|
| [403] |
Tait SW, Green DR (2010) Mitochondria and cell death: outer membrane permeabilization and beyond. Nat Rev Mol Cell Biol 11: 621-632. doi: 10.1038/nrm2952
|
| [404] |
Gross A, McDonnell JM, Korsmeyer SJ (1999) BCL-2 family members and the mitochondria in apoptosis. Genes Dev 13: 1899-1911. doi: 10.1101/gad.13.15.1899
|
| [405] |
Putcha GV, Deshmukh M, Johnson EM (1999) BAX translocation is a critical event in neuronal apoptosis: regulation by neuroprotectants, BCL-2, and caspases. J Neurosci 19: 7476-7485. doi: 10.1523/JNEUROSCI.19-17-07476.1999
|
| [406] |
Edlich F, Banerjee S, Suzuki M, et al. (2011) Bcl-x(L) retrotranslocates Bax from the mitochondria into the cytosol. Cell 145: 104-116. doi: 10.1016/j.cell.2011.02.034
|
| [407] |
Su Y, Ryder J, Li B, et al. (2004) Lithium, a common drug for bipolar disorder treatment, regulates amyloid-beta precursor protein processing. Biochemistry 43: 6899-6908. doi: 10.1021/bi035627j
|
| [408] |
Xu X, Zhang A, Zhu Y, et al. (2018) MFG-E8 reverses microglial-induced neurotoxic astrocyte (A1) via NF-kappaB and PI3K-Akt pathways. J Cell Physiol 234: 904-914. doi: 10.1002/jcp.26918
|
| [409] |
Jimenez S, Torres M, Vizuete M, et al. (2011) Age-dependent accumulation of soluble amyloid beta (Abeta) oligomers reverses the neuroprotective effect of soluble amyloid precursor protein-alpha (sAPP(alpha)) by modulating phosphatidylinositol 3-kinase (PI3K)/Akt-GSK-3beta pathway in Alzheimer mouse model. J Biol Chem 286: 18414-18425. doi: 10.1074/jbc.M110.209718
|
| [410] |
Cho SJ, Yun SM, Jo C, et al. (2019) Altered expression of Notch1 in Alzheimer's disease. PLoS One 14: e0224941. doi: 10.1371/journal.pone.0224941
|
| [411] |
Taylor KL, Henderson AM, Hughes CC (2002) Notch activation during endothelial cell network formation in vitro targets the basic HLH transcription factor HESR-1 and downregulates VEGFR-2/KDR expression. Microvasc Res 64: 372-383. doi: 10.1006/mvre.2002.2443
|
| [412] |
Yoon KJ, Lee HR, Jo YS, et al. (2012) Mind bomb-1 is an essential modulator of long-term memory and synaptic plasticity via the Notch signaling pathway. Mol Brain 5: 40. doi: 10.1186/1756-6606-5-40
|
| [413] |
Basak O, Giachino C, Fiorini E, et al. (2012) Neurogenic subventricular zone stem/progenitor cells are Notch1-dependent in their active but not quiescent state. J Neurosci 32: 5654-5666. doi: 10.1523/JNEUROSCI.0455-12.2012
|
| [414] |
Brai E, Alina Raio N, Alberi L (2016) Notch1 hallmarks fibrillary depositions in sporadic Alzheimer's disease. Acta Neuropathol Commun 4: 64. doi: 10.1186/s40478-016-0327-2
|
| [415] |
Caricasole A, Copani A, Caraci F, et al. (2004) Induction of Dickkopf-1, a negative modulator of the Wnt pathway, is associated with neuronal degeneration in Alzheimer's brain. J Neurosci 24: 6021-6027. doi: 10.1523/JNEUROSCI.1381-04.2004
|
| [416] |
Rosi MC, Luccarini I, Grossi C, et al. (2010) Increased Dickkopf-1 expression in transgenic mouse models of neurodegenerative disease. J Neurochem 112: 1539-1551. doi: 10.1111/j.1471-4159.2009.06566.x
|
| [417] |
Cerpa W, Godoy JA, Alfaro I, et al. (2008) Wnt-7a modulates the synaptic vesicle cycle and synaptic transmission in hippocampal neurons. J Biol Chem 283: 5918-5927. doi: 10.1074/jbc.M705943200
|
| [418] |
Farias GG, Valles AS, Colombres M, et al. (2007) Wnt-7a induces presynaptic colocalization of alpha 7-nicotinic acetylcholine receptors and adenomatous polyposis coli in hippocampal neurons. J Neurosci 27: 5313-5325. doi: 10.1523/JNEUROSCI.3934-06.2007
|
| [419] |
Tapia-Rojas C, Inestrosa NC (2018) Loss of canonical Wnt signaling is involved in the pathogenesis of Alzheimer's disease. Neural Regen Res 13: 1705-1710. doi: 10.4103/1673-5374.238606
|
| [420] |
Elliott C, Rojo AI, Ribe E, et al. (2018) A role for APP in Wnt signalling links synapse loss with beta-amyloid production. Transl Psychiatry 8: 179. doi: 10.1038/s41398-018-0231-6
|
| [421] | Aso E, Ferrer I (2014) Cannabinoids for treatment of Alzheimer's disease: moving toward the clinic. Front Pharmacol 5: 37. |
| [422] |
Solas M, Francis PT, Franco R, et al. (2013) CB2 receptor and amyloid pathology in frontal cortex of Alzheimer's disease patients. Neurobiol Aging 34: 805-808. doi: 10.1016/j.neurobiolaging.2012.06.005
|
| [423] |
Barnado A, Crofford LJ, Oates JC (2016) At the Bedside: Neutrophil extracellular traps (NETs) as targets for biomarkers and therapies in autoimmune diseases. J Leukoc Biol 99: 265-278. doi: 10.1189/jlb.5BT0615-234R
|
| [424] |
Hamilton A, Esseltine JL, DeVries RA, et al. (2014) Metabotropic glutamate receptor 5 knockout reduces cognitive impairment and pathogenesis in a mouse model of Alzheimer's disease. Mol Brain 7: 40. doi: 10.1186/1756-6606-7-40
|
 neurosci-08-01-005-s001.pdf neurosci-08-01-005-s001.pdf |
 |
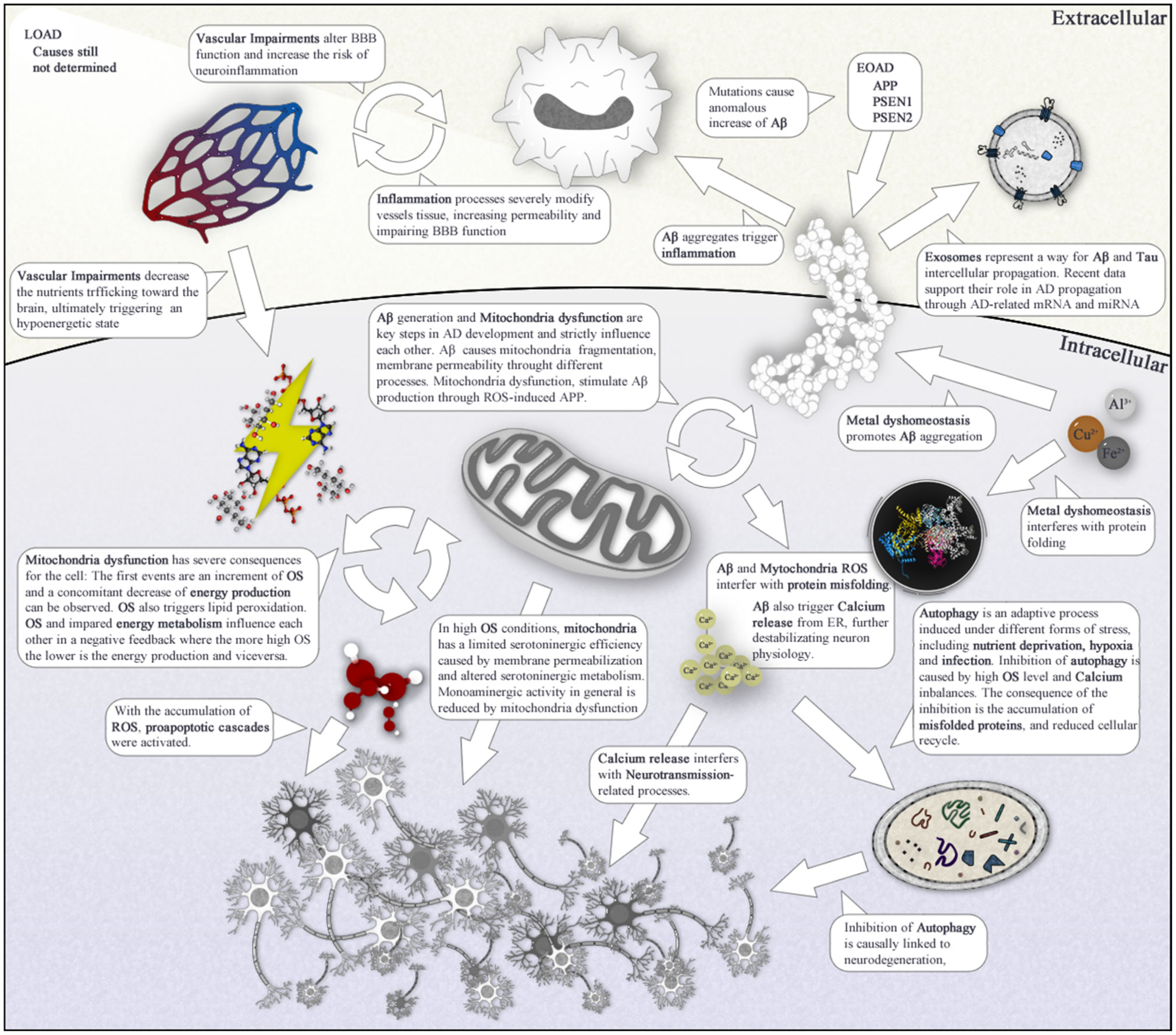
Figures(1) / Tables(1)

Marco Calabrò, Carmela Rinaldi, Giuseppe Santoro, Concetta Crisafulli. The biological pathways of Alzheimer disease: a review[J]. AIMS Neuroscience, 2021, 8(1): 86-132. doi: 10.3934/Neuroscience.2021005







 DownLoad:
DownLoad: