Citation: Peter X. Shaw, Travis Stiles, Christopher Douglas, Daisy Ho, Wei Fan, Hongjun Du, Xu Xiao. Oxidative stress, innate immunity, and age-related macular degeneration[J]. AIMS Molecular Science, 2016, 3(2): 196-221. doi: 10.3934/molsci.2016.2.196

| [1] |
Jager RD, Mieler WF, Miller JW (2008) Age-related macular degeneration. N Engl J Med 358: 2606-2617. doi: 10.1056/NEJMra0801537

|
| [2] | Sunness JS (1999) The natural history of geographic atrophy, the advanced atrophic form of age-related macular degeneration. Mol Vis 5: 25. |
| [3] |
Ying GS, Maguire MG, Complications of Age-related Macular Degeneration Prevention Trial Research G (2011) Development of a risk score for geographic atrophy in complications of the age-related macular degeneration prevention trial. Ophthalmology 118: 332-338. doi: 10.1016/j.ophtha.2010.06.030
|
| [4] | Sarks JP, Sarks SH, Killingsworth MC (1988) Evolution of geographic atrophy of the retinal pigment epithelium. Eye (Lond) 2 (Pt 5): 552-577. |
| [5] |
Ferris FL 3rd, Fine SL, Hyman L (1984) Age-related macular degeneration and blindness due to neovascular maculopathy. Arch Ophthalmol 102: 1640-1642. doi: 10.1001/archopht.1984.01040031330019
|
| [6] |
Chen Y, Zeng J, Zhao C, et al. (2011) Assessing susceptibility to age-related macular degeneration with genetic markers and environmental factors. Arch Ophthalmol 129: 344-351. doi: 10.1001/archophthalmol.2011.10
|
| [7] | American Academy of Ophthalmology. What Are Drusen? 2014. Available from: http://www.aao.org/eye-health/diseases/what-are-drusen |
| [8] |
Shaw PX, Zhang L, Zhang M, et al. (2012) Complement factor H genotypes impact risk of age-related macular degeneration by interaction with oxidized phospholipids. Proc Natl Acad Sci U S A 109: 13757-13762. doi: 10.1073/pnas.1121309109
|
| [9] |
Fritsche LG, Chen W, Schu M, et al. (2013) Seven new loci associated with age-related macular degeneration. Nat Genet 45: 433-439, 439e431-432. doi: 10.1038/ng.2578
|
| [10] |
Beharry S, Zhong M, Molday RS (2004) N-retinylidene-phosphatidylethanolamine is the preferred retinoid substrate for the photoreceptor-specific ABC transporter ABCA4 (ABCR). J Biol Chem 279: 53972-53979. doi: 10.1074/jbc.M405216200
|
| [11] | Allikmets R (2000) Further evidence for an association of ABCR alleles with age-related macular degeneration. The International ABCR Screening Consortium. Am J Hum Genet 67: 487-491. |
| [12] |
Katta S, Kaur I, Chakrabarti S (2009) The molecular genetic basis of age-related macular degeneration: an overview. J Genet 88: 425-449. doi: 10.1007/s12041-009-0064-4
|
| [13] |
Thakkinstian A, Bowe S, McEvoy M, et al. (2006) Association between apolipoprotein E polymorphisms and age-related macular degeneration: A HuGE review and meta-analysis. Am J Epidemiol 164: 813-822. doi: 10.1093/aje/kwj279
|
| [14] |
McKay GJ, Silvestri G, Chakravarthy U, et al. (2011) Variations in apolipoprotein E frequency with age in a pooled analysis of a large group of older people. Am J Epidemiol 173: 1357-1364. doi: 10.1093/aje/kwr015
|
| [15] |
Fritsche LG, Loenhardt T, Janssen A, et al. (2008) Age-related macular degeneration is associated with an unstable ARMS2 (LOC387715) mRNA. Nat Genet 40: 892-896. doi: 10.1038/ng.170
|
| [16] |
Ross RJ, Bojanowski CM, Wang JJ, et al. (2007) The LOC387715 polymorphism and age-related macular degeneration: replication in three case-control samples. Invest Ophthalmol Vis Sci 48: 1128-1132. doi: 10.1167/iovs.06-0999
|
| [17] |
Jakobsdottir J, Conley YP, Weeks DE, et al. (2005) Susceptibility genes for age-related maculopathy on chromosome 10q26. Am J Hum Genet 77: 389-407. doi: 10.1086/444437
|
| [18] |
Dewan A, Liu M, Hartman S, et al. (2006) HTRA1 promoter polymorphism in wet age-related macular degeneration. Science 314: 989-992. doi: 10.1126/science.1133807
|
| [19] |
Yang Z, Camp NJ, Sun H, et al. (2006) A variant of the HTRA1 gene increases susceptibility to age-related macular degeneration. Science 314: 992-993. doi: 10.1126/science.1133811
|
| [20] |
Gold B, Merriam JE, Zernant J, et al. (2006) Variation in factor B (BF) and complement component 2 (C2) genes is associated with age-related macular degeneration. Nat Genet 38: 458-462. doi: 10.1038/ng1750
|
| [21] |
Yates JR, Sepp T, Matharu BK, et al. (2007) Complement C3 variant and the risk of age-related macular degeneration. N Engl J Med 357: 553-561. doi: 10.1056/NEJMoa072618
|
| [22] |
Maller JB, Fagerness JA, Reynolds RC, et al. (2007) Variation in complement factor 3 is associated with risk of age-related macular degeneration. Nat Genet 39: 1200-1201. doi: 10.1038/ng2131
|
| [23] |
Francis PJ, Hamon SC, Ott J, et al. (2009) Polymorphisms in C2, CFB and C3 are associated with progression to advanced age related macular degeneration associated with visual loss. J Med Genet 46: 300-307. doi: 10.1136/jmg.2008.062737
|
| [24] |
Chen W, Stambolian D, Edwards AO, et al. (2010) Genetic variants near TIMP3 and high-density lipoprotein-associated loci influence susceptibility to age-related macular degeneration. Proc Natl Acad Sci U S A 107: 7401-7406. doi: 10.1073/pnas.0912702107
|
| [25] |
Hageman GS, Anderson DH, Johnson LV, et al. (2005) A common haplotype in the complement regulatory gene factor H (HF1/CFH) predisposes individuals to age-related macular degeneration. Proc Natl Acad Sci U S A 102: 7227-7232. doi: 10.1073/pnas.0501536102
|
| [26] |
Wegscheider BJ, Weger M, Renner W, et al. (2007) Association of complement factor H Y402H gene polymorphism with different subtypes of exudative age-related macular degeneration. Ophthalmology 114: 738-742. doi: 10.1016/j.ophtha.2006.07.048
|
| [27] |
Klein RJ, Zeiss C, Chew EY, et al. (2005) Complement factor H polymorphism in age-related macular degeneration. Science 308: 385-389. doi: 10.1126/science.1109557
|
| [28] |
Thakkinstian A, Han P, McEvoy M, et al. (2006) Systematic review and meta-analysis of the association between complement factor H Y402H polymorphisms and age-related macular degeneration. Hum Mol Genet 15: 2784-2790. doi: 10.1093/hmg/ddl220
|
| [29] | Spencer KL, Hauser MA, Olson LM, et al. (2008) Deletion of CFHR3 and CFHR1 genes in age-related macular degeneration. Hum Mol Genet 17: 971-977. |
| [30] |
Raychaudhuri S, Ripke S, Li M, et al. (2010) Associations of CFHR1–CFHR3 deletion and a CFH SNP to age-related macular degeneration are not independent. Nat Genet 42: 553-556. doi: 10.1038/ng0710-553
|
| [31] | Fagerness JA, Maller JB, Neale BM, et al. (2008) Variation near complement factor I is associated with risk of advanced AMD. Eur J Hum Genet 17: 100-104. |
| [32] |
Combadière C, Feumi C, Raoul W, et al. (2007) CX3CR1-dependent subretinal microglia cell accumulation is associated with cardinal features of age-related macular degeneration. J Clin Investig 117: 2920-2928. doi: 10.1172/JCI31692
|
| [33] | Tuo J, Smith BC, Bojanowski CM, et al. (2004) The involvement of sequence variation and expression of CX3CR1 in the pathogenesis of age-related macular degeneration. FASEB J 18: 1297-1299. |
| [34] |
Tuo J, Ning B, Bojanowski CM, et al. (2006) Synergic effect of polymorphisms in ERCC6 5′ flanking region and complement factor H on age-related macular degeneration predisposition. Proc Natl Acad Sci U S A 103: 9256-9261. doi: 10.1073/pnas.0603485103
|
| [35] |
Neale BM, Fagerness J, Reynolds R, et al. (2010) Genome-wide association study of advanced age-related macular degeneration identifies a role of the hepatic lipase gene (LIPC). Proc Natl Acad Sci U S A 107: 7395-7400. doi: 10.1073/pnas.0912019107
|
| [36] |
Yu Y, Reynolds R, Fagerness J, et al. (2011) Association of variants in the LIPC and ABCA1 genes with intermediate and large drusen and advanced age-related macular degeneration. Invest Ophthalmol Vis Sci 52: 4663-4670. doi: 10.1167/iovs.10-7070
|
| [37] |
Ardeljan D, Meyerle CB, Agron E, et al. (2013) Influence of TIMP3/SYN3 polymorphisms on the phenotypic presentation of age-related macular degeneration. Eur J Hum Genet 21: 1152-1157. doi: 10.1038/ejhg.2013.14
|
| [38] |
Chen W, Stambolian D, Edwards AO, et al. (2010) Genetic variants near TIMP3 and high-density lipoprotein-associated loci influence susceptibility to age-related macular degeneration. Proc Natl Acad Sci U S A 107: 7401-7406. doi: 10.1073/pnas.0912702107
|
| [39] |
Kaur I, Rathi S, Chakrabarti S (2010) Variations in TIMP3 are associated with age-related macular degeneration. Proc Natl Acad Sci U S A 107: E112-E113. doi: 10.1073/pnas.1007476107
|
| [40] |
Yang Z, Stratton C, Francis PJ, et al. (2008) Toll-like receptor 3 and geographic atrophy in age-related macular degeneration. N Engl J Med 359: 1456-1463. doi: 10.1056/NEJMoa0802437
|
| [41] |
Edwards AO, Chen D, Fridley BL, et al. (2008) Toll-like receptor polymorphisms and age-related macular degeneration. Invest Ophthalmol Vis Sci 49: 1652-1659. doi: 10.1167/iovs.07-1378
|
| [42] |
Ferwerda B, McCall MB, Alonso S, et al. (2007) TLR4 polymorphisms, infectious diseases, and evolutionary pressure during migration of modern humans. Proc Natl Acad Sci U S A 104: 16645-16650. doi: 10.1073/pnas.0704828104
|
| [43] |
Zareparsi S, Buraczynska M, Branham KE, et al. (2005) Toll-like receptor 4 variant D299G is associated with susceptibility to age-related macular degeneration. Hum Mol Genet 14: 1449-1455. doi: 10.1093/hmg/ddi154
|
| [44] |
Galan A, Ferlin A, Caretti L, et al. (2010) Association of age-related macular degeneration with polymorphisms in vascular endothelial growth factor and its receptor. Ophthalmology 117: 1769-1774. doi: 10.1016/j.ophtha.2010.01.030
|
| [45] | Fang AM, Lee AY, Kulkarni M, et al. (2009) Polymorphisms in the VEGFA and VEGFR-2 genes and neovascular age-related macular degeneration. Mol Vis 15: 2710-2719. |
| [46] |
Churchill AJ, Carter JG, Ramsden C, et al. (2008) VEGF polymorphisms are associated with severity of diabetic retinopathy. Invest Ophthalmol Vis Sci 49: 3611-3616. doi: 10.1167/iovs.07-1383
|
| [47] |
Jozsi M, Zipfel PF (2008) Factor H family proteins and human diseases. Trends Immunol 29: 380-387. doi: 10.1016/j.it.2008.04.008
|
| [48] |
Fritsche LG, Lauer N, Hartmann A, et al. (2010) An imbalance of human complement regulatory proteins CFHR1, CFHR3 and factor H influences risk for age-related macular degeneration (AMD). Hum Mol Genet 19: 4694-4704. doi: 10.1093/hmg/ddq399
|
| [49] |
Zipfel PF, Edey M, Heinen S, et al. (2007) Deletion of complement factor H-related genes CFHR1 and CFHR3 is associated with atypical hemolytic uremic syndrome. PLoS Genet 3: e41. doi: 10.1371/journal.pgen.0030041
|
| [50] | Zipfel PF, Skerka C (2009) Complement regulators and inhibitory proteins. Nat Rev Immunol 9: 729-740. |
| [51] |
Ricklin D, Hajishengallis G, Yang K, et al. (2010) Complement: a key system for immune surveillance and homeostasis. Nat Immunol 11: 785-797. doi: 10.1038/ni.1923
|
| [52] |
Hughes AE, Orr N, Esfandiary H, et al. (2006) A common CFH haplotype, with deletion of CFHR1 and CFHR3, is associated with lower risk of age-related macular degeneration. Nat Genet 38: 1173-1177. doi: 10.1038/ng1890
|
| [53] |
Zhao J, Wu H, Khosravi M, et al. (2011) Association of genetic variants in complement factor H and factor H-related genes with systemic lupus erythematosus susceptibility. PLoS Genet 7: e1002079. doi: 10.1371/journal.pgen.1002079
|
| [54] |
Tortajada A, Yebenes H, Abarrategui-Garrido C, et al. (2013) C3 glomerulopathy-associated CFHR1 mutation alters FHR oligomerization and complement regulation. J Clin Invest 123: 2434-2446. doi: 10.1172/JCI68280
|
| [55] |
Zipfel PF, Mache C, Muller D, et al. (2010) DEAP-HUS: deficiency of CFHR plasma proteins and autoantibody-positive form of hemolytic uremic syndrome. Pediatr Nephrol 25: 2009-2019. doi: 10.1007/s00467-010-1446-9
|
| [56] | Jozsi M, Licht C, Strobel S, et al. (2008) Factor H autoantibodies in atypical hemolytic uremic syndrome correlate with CFHR1/CFHR3 deficiency. Blood 111: 1512-1514. |
| [57] |
Dragon-Durey MA, Blanc C, Marliot F, et al. (2009) The high frequency of complement factor H related CFHR1 gene deletion is restricted to specific subgroups of patients with atypical haemolytic uraemic syndrome. J Med Genet 46: 447-450. doi: 10.1136/jmg.2008.064766
|
| [58] |
Kishan AU, Modjtahedi BS, Martins EN, et al. (2011) Lipids and age-related macular degeneration. Surv Ophthalmol 56: 195-213. doi: 10.1016/j.survophthal.2010.08.008
|
| [59] |
Klein R, Myers CE, Buitendijk GH, et al. (2014) Lipids, lipid genes, and incident age-related macular degeneration: the three continent age-related macular degeneration consortium. Am J Ophthalmol 158: 513-524 e513. doi: 10.1016/j.ajo.2014.05.027
|
| [60] |
Martiskainen H, Haapasalo A, Kurkinen KM, et al. (2013) Targeting ApoE4/ApoE receptor LRP1 in Alzheimer's disease. Expert Opin Ther Targets 17: 781-794. doi: 10.1517/14728222.2013.789862
|
| [61] |
Liu CC, Kanekiyo T, Xu H, et al. (2013) Apolipoprotein E and Alzheimer disease: risk, mechanisms and therapy. Nat Rev Neurol 9: 106-118. doi: 10.1038/nrneurol.2012.263
|
| [62] |
Kehoe P, Wavrant-De Vrieze F, Crook R, et al. (1999) A full genome scan for late onset Alzheimer's disease. Hum Mol Genet 8: 237-245. doi: 10.1093/hmg/8.2.237
|
| [63] |
Cezario SM, Calastri MC, Oliveira CI, et al. (2015) Association of high-density lipoprotein and apolipoprotein E genetic variants with age-related macular degeneration. Arq Bras Oftalmol 78: 85-88. doi: 10.5935/0004-2749.20150023
|
| [64] |
Levy O, Calippe B, Lavalette S, et al. (2015) Apolipoprotein E promotes subretinal mononuclear phagocyte survival and chronic inflammation in age-related macular degeneration. EMBO Mol Med 7: 211-226. doi: 10.15252/emmm.201404524
|
| [65] |
Zhang L, Lim SL, Du H, et al. (2012) High temperature requirement factor A1 (HTRA1) gene regulates angiogenesis through transforming growth factor-beta family member growth differentiation factor 6. J Biol Chem 287: 1520-1526. doi: 10.1074/jbc.M111.275990
|
| [66] | Langton KP, McKie N, Curtis A, et al. (2000) A novel tissue inhibitor of metalloproteinases-3 mutation reveals a common molecular phenotype in Sorsby's fundus dystrophy. J Biol Chem 275: 27027-27031. |
| [67] |
Clarke M, Mitchell KW, Goodship J, et al. (2001) Clinical features of a novel TIMP-3 mutation causing Sorsby's fundus dystrophy: implications for disease mechanism. Br J Ophthalmol 85: 1429-1431. doi: 10.1136/bjo.85.12.1429
|
| [68] | Fritsche LG, Igl W, Bailey JN, et al. (2016) A large genome-wide association study of age-related macular degeneration highlights contributions of rare and common variants. Nat Genet 48: 134-143. |
| [69] |
AREDS (2001) A randomized, placebo-controlled, clinical trial of high-dose supplementation with vitamins C and E, beta carotene, and zinc for age-related macular degeneration and vision loss: AREDS report no. 8. Arch Ophthalmol 119: 1417-1436. doi: 10.1001/archopht.119.10.1417
|
| [70] |
Curcio CA, Johnson M, Huang JD, et al. (2009) Aging, age-related macular degeneration, and the response-to-retention of apolipoprotein B-containing lipoproteins. Prog Retin Eye Res 28: 393-422. doi: 10.1016/j.preteyeres.2009.08.001
|
| [71] | Hawkins BS, Bird A, Klein R, et al. (1999) Epidemiology of age-related macular degeneration. Mol Vis 5: 26. |
| [72] | Khotcharrat R, Patikulsila D, Hanutsaha P, et al. (2015) Epidemiology of Age-Related Macular Degeneration among the Elderly Population in Thailand. J Med Assoc Thai 98: 790-797. |
| [73] |
Kiernan DF, Hariprasad SM, Rusu IM, et al. (2010) Epidemiology of the association between anticoagulants and intraocular hemorrhage in patients with neovascular age-related macular degeneration. Retina 30: 1573-1578. doi: 10.1097/IAE.0b013e3181e2266d
|
| [74] |
Klein R, Peto T, Bird A, et al. (2004) The epidemiology of age-related macular degeneration. Am J Ophthalmol 137: 486-495. doi: 10.1016/j.ajo.2003.11.069
|
| [75] |
Meyers KJ, Liu Z, Millen AE, et al. (2015) Joint Associations of Diet, Lifestyle, and Genes with Age-Related Macular Degeneration. Ophthalmology 122: 2286-2294. doi: 10.1016/j.ophtha.2015.07.029
|
| [76] |
Seddon JM, George S, Rosner B (2006) Cigarette smoking, fish consumption, omega-3 fatty acid intake, and associations with age-related macular degeneration: the US Twin Study of Age-Related Macular Degeneration. Arch Ophthalmol 124: 995-1001. doi: 10.1001/archopht.124.7.995
|
| [77] |
Schick T, Ersoy L, Lechanteur YT, et al. (2016) History of Sunlight Exposure Is a Risk Factor for Age-Related Macular Degeneration. Retina 36: 787-790. doi: 10.1097/IAE.0000000000000756
|
| [78] |
Millen AE, Meyers KJ, Liu Z, et al. (2015) Association between vitamin D status and age-related macular degeneration by genetic risk. JAMA Ophthalmol 133: 1171-1179. doi: 10.1001/jamaophthalmol.2015.2715
|
| [79] |
Binder CJ, Chang MK, Shaw PX, et al. (2002) Innate and acquired immunity in atherogenesis. Nat Med 8: 1218-1226. doi: 10.1038/nm1102-1218
|
| [80] | Brewer GJ (2007) Iron and copper toxicity in diseases of aging, particularly atherosclerosis and Alzheimer's disease. Exp Biol Med (Maywood) 232: 323-335. |
| [81] |
Beatty S, Koh H, Phil M, et al. (2000) The role of oxidative stress in the pathogenesis of age-related macular degeneration. Surv Ophthalmol 45: 115-134. doi: 10.1016/S0039-6257(00)00140-5
|
| [82] |
Hollyfield JG, Bonilha VL, Rayborn ME, et al. (2008) Oxidative damage-induced inflammation initiates age-related macular degeneration. Nat Med 14: 194-198. doi: 10.1038/nm1709
|
| [83] |
Kim GH, Kim JE, Rhie SJ, et al. (2015) The Role of Oxidative Stress in Neurodegenerative Diseases. Exp Neurobiol 24: 325-340. doi: 10.5607/en.2015.24.4.325
|
| [84] |
Smith W, Assink J, Klein R, et al. (2001) Risk factors for age-related macular degeneration: Pooled findings from three continents. Ophthalmology 108: 697-704. doi: 10.1016/S0161-6420(00)00580-7
|
| [85] |
Damian J, Pastor R, Armada F, et al. (2006) Epidemiology of age-related macular degeneration. Situation in Spain. Aten Primaria 38: 51-57. doi: 10.1157/13090016
|
| [86] | Wang YQ, Dong XG (2005) Progress in the study of epidemiology and etiology of age-related macular degeneration. Zhonghua Yan Ke Za Zhi 41: 377-381. |
| [87] |
Bonastre J, Le Pen C, Anderson P, et al. (2002) The epidemiology, economics and quality of life burden of age-related macular degeneration in France, Germany, Italy and the United Kingdom. Eur J Health Econ 3: 94-102. doi: 10.1007/s10198-002-0104-y
|
| [88] | Seddon JM, Chen CA (2004) The epidemiology of age-related macular degeneration. Int Ophthalmol Clin 44: 17-39. |
| [89] |
Friedman DS, O'Colmain BJ, Munoz B, et al. (2004) Prevalence of age-related macular degeneration in the United States. Arch Ophthalmol 122: 564-572. doi: 10.1001/archopht.122.4.564
|
| [90] |
Solberg Y, Rosner M, Belkin M (1998) The association between cigarette smoking and ocular diseases. Surv Ophthalmol 42: 535-547. doi: 10.1016/S0039-6257(98)00002-2
|
| [91] | Shen JK, Dong A, Hackett SF, et al. (2007) Oxidative damage in age-related macular degeneration. Histol Histopathol 22: 1301-1308. |
| [92] |
Thurman JM, Renner B, Kunchithapautham K, et al. (2009) Oxidative stress renders retinal pigment epithelial cells susceptible to complement-mediated injury. J Biol Chem 284: 16939-16947. doi: 10.1074/jbc.M808166200
|
| [93] |
Wu Z, Lauer TW, Sick A, et al. (2007) Oxidative stress modulates complement factor H expression in retinal pigmented epithelial cells by acetylation of FOXO3. J Biol Chem 282: 22414-22425. doi: 10.1074/jbc.M702321200
|
| [94] | Catala A (2011) Lipid peroxidation of membrane phospholipids in the vertebrate retina. Front Biosci (Schol Ed) 3: 52-60. |
| [95] |
Kevany BM, Palczewski K (2010) Phagocytosis of retinal rod and cone photoreceptors. Physiology (Bethesda) 25: 8-15. doi: 10.1152/physiol.00038.2009
|
| [96] |
Shaw PX, Horkko S, Chang MK, et al. (2000) Natural antibodies with the T15 idiotype may act in atherosclerosis, apoptotic clearance, and protective immunity. J Clin Invest 105: 1731-1740. doi: 10.1172/JCI8472
|
| [97] |
Jeitner TM, Voloshyna I, Reiss AB (2011) Oxysterol derivatives of cholesterol in neurodegenerative disorders. Curr Med Chem 18: 1515-1525. doi: 10.2174/092986711795328445
|
| [98] |
Larrayoz IM, Huang JD, Lee JW, et al. (2010) 7-ketocholesterol-induced inflammation: involvement of multiple kinase signaling pathways via NFkappaB but independently of reactive oxygen species formation. Invest Ophthalmol Vis Sci 51: 4942-4955. doi: 10.1167/iovs.09-4854
|
| [99] |
Joffre C, Leclere L, Buteau B, et al. (2007) Oxysterols induced inflammation and oxidation in primary porcine retinal pigment epithelial cells. Curr Eye Res 32: 271-280. doi: 10.1080/02713680601187951
|
| [100] | Mylonas C, Kouretas D (1999) Lipid peroxidation and tissue damage. In Vivo 13: 295-309. |
| [101] |
Duryee MJ, Klassen LW, Schaffert CS, et al. (2010) Malondialdehyde-acetaldehyde adduct is the dominant epitope after MDA modification of proteins in atherosclerosis. Free Radic Biol Med 49: 1480-1486. doi: 10.1016/j.freeradbiomed.2010.08.001
|
| [102] | Kaji Y, Usui T, Oshika T, et al. (2000) Advanced glycation end products in diabetic corneas. Invest Ophthalmol Vis Sci 41: 362-368. |
| [103] | Handa JT, Verzijl N, Matsunaga H, et al. (1999) Increase in the advanced glycation end product pentosidine in Bruch's membrane with age. Invest Ophthalmol Vis Sci 40: 775-779. |
| [104] |
Renganathan K, Ebrahem Q, Vasanji A, et al. (2008) Carboxyethylpyrrole adducts, age-related macular degeneration and neovascularization. Adv Exp Med Biol 613: 261-267. doi: 10.1007/978-0-387-74904-4_30
|
| [105] | Barja G, Herrero A (2000) Oxidative damage to mitochondrial DNA is inversely related to maximum life span in the heart and brain of mammals. FASEB J 14: 312-318. |
| [106] |
Dib B, Lin H, Maidana DE, et al. (2015) Mitochondrial DNA has a pro-inflammatory role in AMD. Biochim Biophys Acta 1853: 2897-2906. doi: 10.1016/j.bbamcr.2015.08.012
|
| [107] |
Lin H, Xu H, Liang FQ, et al. (2011) Mitochondrial DNA damage and repair in RPE associated with aging and age-related macular degeneration. Invest Ophthalmol Vis Sci 52: 3521-3529. doi: 10.1167/iovs.10-6163
|
| [108] | Wang AL, Lukas TJ, Yuan M, et al. (2008) Increased mitochondrial DNA damage and down-regulation of DNA repair enzymes in aged rodent retinal pigment epithelium and choroid. Mol Vis 14: 644-651. |
| [109] |
Terluk MR, Kapphahn RJ, Soukup LM, et al. (2015) Investigating mitochondria as a target for treating age-related macular degeneration. J Neurosci 35: 7304-7311. doi: 10.1523/JNEUROSCI.0190-15.2015
|
| [110] |
Perez VL, Caspi RR (2015) Immune mechanisms in inflammatory and degenerative eye disease. Trends Immunol 36: 354-363. doi: 10.1016/j.it.2015.04.003
|
| [111] |
Du H, Sun X, Guma M, et al. (2013) JNK inhibition reduces apoptosis and neovascularization in a murine model of age-related macular degeneration. Proc Natl Acad Sci U S A 110: 2377-2382. doi: 10.1073/pnas.1221729110
|
| [112] |
Horkko S, Binder CJ, Shaw PX, et al. (2000) Immunological responses to oxidized LDL. Free Radic Biol Med 28: 1771-1779. doi: 10.1016/S0891-5849(00)00333-6
|
| [113] | Berliner JA, Navab M, Fogelman AM, et al. (1995) Atherosclerosis: basic mechanisms. Oxidation, inflammation, and genetics. Circulation 91: 2488-2496. |
| [114] | Ebrahimi KB, Handa JT (2011) Lipids, lipoproteins, and age-related macular degeneration. J Lipids 2011: 802059. |
| [115] |
Chou MY, Hartvigsen K, Hansen LF, et al. (2008) Oxidation-specific epitopes are important targets of innate immunity. J Intern Med 263: 479-488. doi: 10.1111/j.1365-2796.2008.01968.x
|
| [116] |
Weismann D, Hartvigsen K, Lauer N, et al. (2011) Complement factor H binds malondialdehyde epitopes and protects from oxidative stress. Nature 478: 76-81. doi: 10.1038/nature10449
|
| [117] | Ambati J, Atkinson JP, Gelfand BD (2013) Immunology of age-related macular degeneration. Nat Rev Immunol 13: 438-451. |
| [118] |
de Oliveira Dias JR, Rodrigues EB, Maia M, et al. (2011) Cytokines in neovascular age-related macular degeneration: fundamentals of targeted combination therapy. BrJ Ophthalmol 95: 1631-1637. doi: 10.1136/bjo.2010.186361
|
| [119] |
Mo FM, Proia AD, Johnson WH, et al. (2010) Interferon gamma-inducible protein-10 (IP-10) and eotaxin as biomarkers in age-related macular degeneration. Invest Ophthalmol Vis Sci 51: 4226-4236. doi: 10.1167/iovs.09-3910
|
| [120] | Wang JC, Cao S, Wang A, et al. (2015) CFH Y402H polymorphism is associated with elevated vitreal GM-CSF and choroidal macrophages in the postmortem human eye. Mol Vis 21: 264-272. |
| [121] |
Lad EM, Cousins SW, Van Arnam JS, et al. (2015) Abundance of infiltrating CD163+ cells in the retina of postmortem eyes with dry and neovascular age-related macular degeneration. Graefes Arch Clin Exp Ophthalmol 253: 1941-1945. doi: 10.1007/s00417-015-3094-z
|
| [122] |
Doyle SL, Campbell M, Ozaki E, et al. (2012) NLRP3 has a protective role in age-related macular degeneration through the induction of IL-18 by drusen components. Nat Med 18: 791-798. doi: 10.1038/nm.2717
|
| [123] |
Doyle SL, Lopez FJ, Celkova L, et al. (2015) IL-18 Immunotherapy for Neovascular AMD: Tolerability and Efficacy in Nonhuman Primates. Invest Ophthalmol Vis Sci 56: 5424-5430. doi: 10.1167/iovs.15-17264
|
| [124] | Doyle SL, Ozaki E, Brennan K, et al. (2014) IL-18 attenuates experimental choroidal neovascularization as a potential therapy for wet age-related macular degeneration. Sci Transl Med 6: 230ra244. |
| [125] | Skerka C, Zipfel PF (2008) Complement factor H related proteins in immune diseases. Vaccine 26 Suppl 8: I9-14. |
| [126] |
Kelly J, Ali Khan A, Yin J, et al. (2007) Senescence regulates macrophage activation and angiogenic fate at sites of tissue injury in mice. J Clin Invest 117: 3421-3426. doi: 10.1172/JCI32430
|
| [127] |
Xu H, Chen M, Forrester JV (2009) Para-inflammation in the aging retina. Prog Retin Eye Res 28: 348-368. doi: 10.1016/j.preteyeres.2009.06.001
|
| [128] |
Anderson DH, Radeke MJ, Gallo NB, et al. (2010) The pivotal role of the complement system in aging and age-related macular degeneration: hypothesis re-visited. Prog Retin Eye Res 29: 95-112. doi: 10.1016/j.preteyeres.2009.11.003
|
| [129] |
Crabb JW, Miyagi M, Gu X, et al. (2002) Drusen proteome analysis: an approach to the etiology of age-related macular degeneration. Proc Natl Acad Sci U S A 99: 14682-14687. doi: 10.1073/pnas.222551899
|
| [130] |
Yuan X, Gu X, Crabb JS, et al. (2010) Quantitative proteomics: comparison of the macular Bruch membrane/choroid complex from age-related macular degeneration and normal eyes. Mol Cell Proteomics 9: 1031-1046. doi: 10.1074/mcp.M900523-MCP200
|
| [131] |
Johnson LV, Ozaki S, Staples MK, et al. (2000) A potential role for immune complex pathogenesis in drusen formation. Exp Eye Res 70: 441-449. doi: 10.1006/exer.1999.0798
|
| [132] |
Wang L, Clark ME, Crossman DK, et al. (2010) Abundant lipid and protein components of drusen. PLoS One 5: e10329. doi: 10.1371/journal.pone.0010329
|
| [133] |
Vierkotten S, Muether PS, Fauser S (2011) Overexpression of HTRA1 leads to ultrastructural changes in the elastic layer of Bruch's membrane via cleavage of extracellular matrix components. PLoS One 6: e22959. doi: 10.1371/journal.pone.0022959
|
| [134] |
Skeie J, Mullins R (2009) Macrophages in neovascular age-related macular degeneration: friends or foes? Eye 23: 747-755. doi: 10.1038/eye.2008.206
|
| [135] |
Killingsworth M, Sarks J, Sarks S (1990) Macrophages related to Bruch’s membrane in age-related macular degeneration. Eye 4: 613-621. doi: 10.1038/eye.1990.86
|
| [136] |
Combadiere C, Feumi C, Raoul W, et al. (2007) CX3CR1-dependent subretinal microglia cell accumulation is associated with cardinal features of age-related macular degeneration. J Clin Investig 117: 2920-2928. doi: 10.1172/JCI31692
|
| [137] |
Liang KJ, Lee JE, Wang YD, et al. (2009) Regulation of dynamic behavior of retinal microglia by CX3CR1 signaling. Invest Ophthalmol Vis Sci 50: 4444-4451. doi: 10.1167/iovs.08-3357
|
| [138] | Grossniklaus H, Ling J, Wallace T, et al. (2002) Macrophage and retinal pigment epithelium expression of angiogenic cytokines in choroidal neovascularization. Mol Vis 8: 119-126. |
| [139] |
Yamada K, Sakurai E, Itaya M, et al. (2007) Inhibition of Laser-Induced Choroidal Neovascularization by Atorvastatin by Downregulation of Monocyte Chemotactic Protein-1 Synthesis in Mice. Invest Ophthalmol Vis Sci 48: 1839-1843. doi: 10.1167/iovs.06-1085
|
| [140] | Mettu PS, Wielgus AR, Ong SS, et al. (2012) Retinal pigment epithelium response to oxidant injury in the pathogenesis of early age-related macular degeneration. Mol Asp Med 33: 376-398. |
| [141] |
Sica A, Mantovani A (2012) Macrophage plasticity and polarization: in vivo veritas. J Clin Investig 122: 787-795. doi: 10.1172/JCI59643
|
| [142] |
Cherepanoff S, McMenamin P, Gillies MC, et al. (2010) Bruch's membrane and choroidal macrophages in early and advanced age-related macular degeneration. Br J Ophthalmol 94: 918-925. doi: 10.1136/bjo.2009.165563
|
| [143] | Sindrilaru A, Peters T, Wieschalka S, et al. An unrestrained proinflammatory M1 macrophage population induced by iron impairs wound healing in humans and mice. J Clin Investig 121: 985-997. |
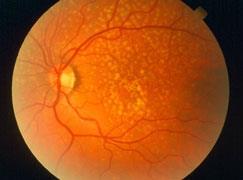
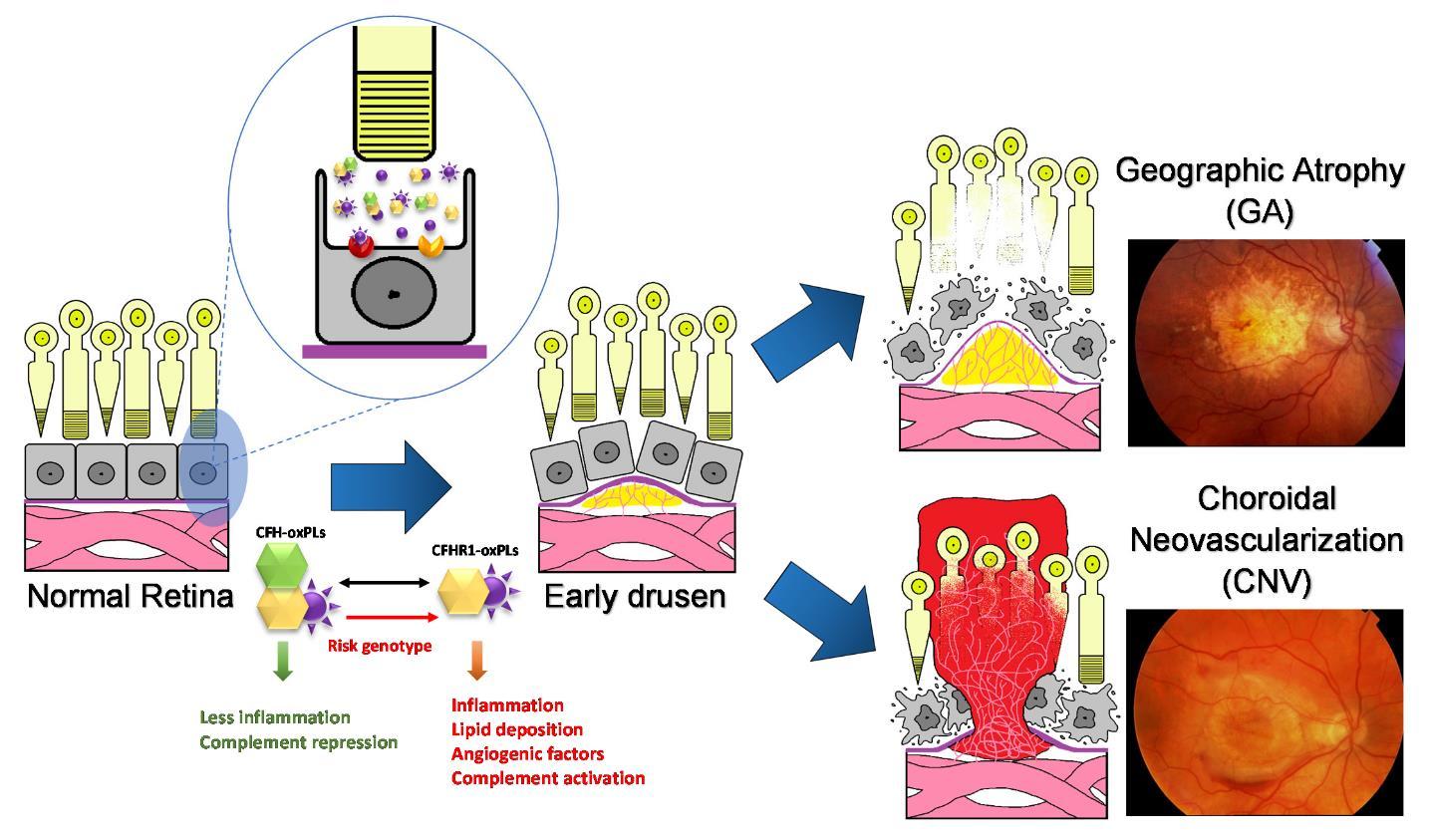
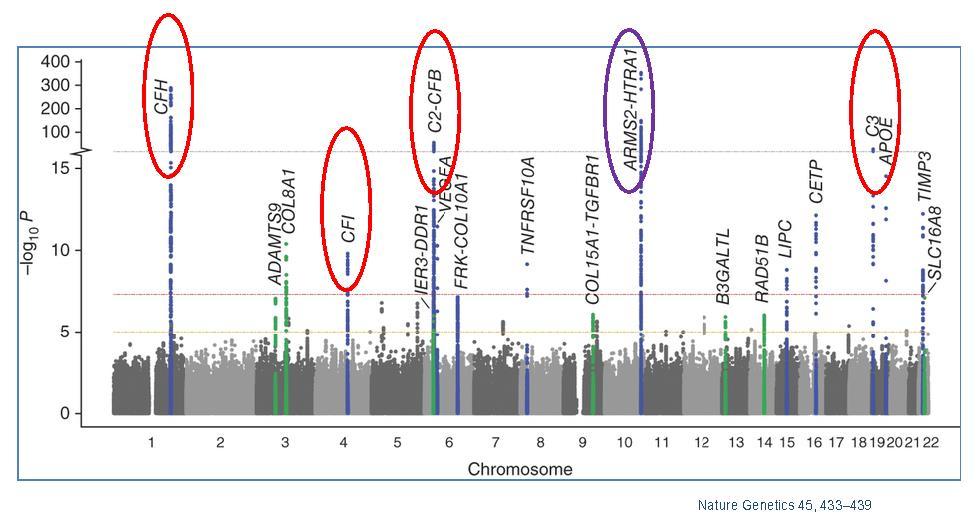

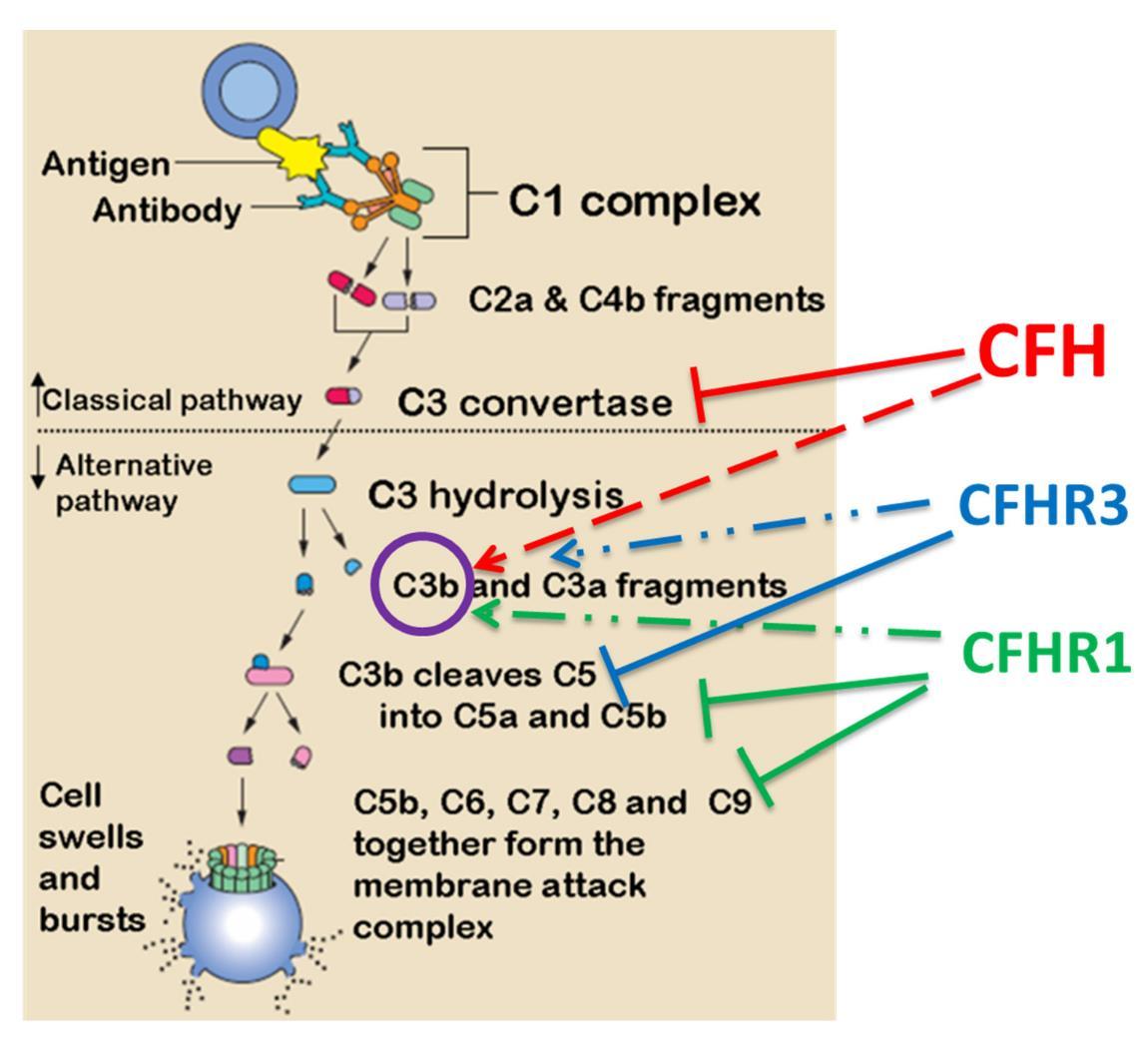
Figures(7) / Tables(1)

Peter X. Shaw, Travis Stiles, Christopher Douglas, Daisy Ho, Wei Fan, Hongjun Du, Xu Xiao. Oxidative stress, innate immunity, and age-related macular degeneration[J]. AIMS Molecular Science, 2016, 3(2): 196-221. doi: 10.3934/molsci.2016.2.196







 DownLoad:
DownLoad: