Citation: Satarupa Dey, Amal Kanti Paul. Assessment of heavy metal tolerance and hexavalent chromium reducing potential of Corynebacterium paurometabolum SKPD 1204 isolated from chromite mine seepage[J]. AIMS Bioengineering, 2016, 3(3): 337-351. doi: 10.3934/bioeng.2016.3.337

| [1] | Mondal SK (2009) Chromite and PGE Deposits of Mesoarchaean Ultramafic-Mafic Suites within the Greenstone Belts of the Singhbhum Craton,India: Implications for Mantle Heterogeneity and Tectonic Setting. J Geo SocIndia 73: 36–51. |
| [2] |
Godgul G, Sahu KC (1995) Chromium contamination from chromium mine. Environ Geol 25: 251–257. doi: 10.1007/BF00766754

|
| [3] | Gibb HJ, Lee PS, Pinsky PF, et al. (2000a) Lung cancer among workers in chromium chemical production. Am J Indus Med 38: 115–126. |
| [4] | Gibb HJ, Lee PS, Pinsky PF, et al. (2000b) Clinical findings of irritation among chromium chemical production workers. Am J Indus Med 38: 127–131. |
| [5] |
Fiol N, Escudero C, Villaescusa I (2008) Chromium sorption and Cr(VI) reduction to Cr(III) by Grape Stalks and Yohimbe Bark. Biores Technol 99: 5030–5036. doi: 10.1016/j.biortech.2007.09.007
|
| [6] |
Cheung KH, Gu JD (2003) Reduction of chromate (CrO42-) by an enrichment consortium and an isolate of marine sulfate-reducing bacteria. Chemosphere 52: 1523–1529. doi: 10.1016/S0045-6535(03)00491-0
|
| [7] | Dey S, Paul AK, (2012) Optimization of cultural conditions for growth associated chromate reduction by Arthrobacter sp. SUK 1201 isolated from chromite mine overburden. J Haz Mat 213–214: 200–206. |
| [8] | Dhal B, Thatoi HN, Das NN et al. (2010) Reduction of hexavalent chromium by Bacillus sp. isolated from chromite mine soils and characterization of reduced product. J Chem Technol Biotechnol 85: 1471–1479. |
| [9] |
Samuel J, Paul ML, Pulimi M et al. (2012) Hexavalent chromium bioremoval through adaptation and consortia development from sukinda chromite mine isolates. Ind Eng Chem Res 51: 3740–3749. doi: 10.1021/ie201796s
|
| [10] |
Wang YT, Xiao C (1995) Factors affecting hexavalent chromium reduction in pure cultures of bacteria. Water Res 29: 2467–2474. doi: 10.1016/0043-1354(95)00093-Z
|
| [11] | Park CH, Keyhan B, Wielinga B et al. (2000) Purification to homogeneity and characterization of a novel Pseudomonas putida chromate reductase, Appl Environ Microbiol 66: 1788–1795. |
| [12] | Lowry OH, Rosebrough NJ, Farr AL et al. (1951) Protein measurements with the folin phenol reagent. J Biol Chem 193: 265–275. |
| [13] | Calomiris JJ, Armstrong TL, Seidler RJ (1984) Association of metal-tolerance with multiple antibiotic resistance of bacteria isolated from drinking water, Appl Environ Microbiol 47: 1238–1242 |
| [14] |
Camargo FAO, Okeke BC, Bento FM, et al. (2003) Chromate reduction by chromium-resistant bacteria isolated from soils contaminated with dichromate. J Environ Qual 32: 1228–1233. doi: 10.2134/jeq2003.1228
|
| [15] |
Dey S, Paul AK, (2010) Occurrence and evaluation of chromium reducing bacteria in seepage water from chromite mine quarries of Orissa,India. J Water Res Protect 2: 380–388. doi: 10.4236/jwarp.2010.24044
|
| [16] | Dey S, Pandit B, Paul AK (2014). Reduction of hexavalent chromium by viable whole cells of chromium resistant bacteria isolated from chromite mining environment. J Min 2014: 941341. |
| [17] | Steinhaus EA (1941) A study of the bacteria associated with thirty species of insects. J Bacteriol 42: 757–790. |
| [18] | Viti P, Pace A, Giovannetti L (2003) Characterisation of chromium-resistant bacteria isolated from chromium-contaminated soil by tannery activity Curr Microbiol 46: 1–5. |
| [19] |
Tahri Joutey N, Bahafid W, Sayel H, et al. (2014) Hexavalent chromium removal by a novel Serratia proteamaculans isolated from the bank of Sebou River (Morocco). Environ Sci Pollut Res 21: 3060–3072 doi: 10.1007/s11356-013-2249-x
|
| [20] |
Kang C, Wu P, Li Y, et al. (2014) Estimates of heavy metal tolerance and chromium(VI) reducing ability of Pseudomonas aeruginosa CCTCC AB93066: chromium(VI) toxicity and environmental parameters optimization. World J Microbiol Biotechnol 30: 2733–2746 doi: 10.1007/s11274-014-1697-x
|
| [21] | Rehman A, Zahoor A, Muneer B, et al. (2008) Chromium tolerance and reduction potential of a Bacillus sp.ev3 isolated from metal contaminated wastewater. Bull Environ Cont Toxicol 81: 25–29. |
| [22] |
Branco R, Chung AP, Johnston T, et al. (2008) The chromate-inducible chrBACF operon from the transposable element TnOtChr confers resistance to chromium(VI) and superoxide. J Bacteriol 190: 6996–7003. doi: 10.1128/JB.00289-08
|
| [23] |
Pal A, Paul AK (2004) Aerobic chromate reduction by chromium resistant bacteria isolated from serpentine soil. Microbiol Res 159: 347–354. doi: 10.1016/j.micres.2004.08.001
|
| [24] | Jain PK, Ramachandran S, Shukla V, et al. (2009) Characterization of metal and antibiotic resistance in a bacterial population isolated from copper mining industry. Int J Integ Biol 6: 57–61. |
| [25] |
Poopal AC, Laxman RS (2009) Studies on biological reduction of chromate by Streptomyces griseus. J Haz Mat 169: 539–545. doi: 10.1016/j.jhazmat.2009.03.126
|
| [26] |
Das S, Mishra J, Das SK, et al (2014) Investigation on mechanism of Cr(VI) reduction and removal by Bacillus amyloliquefaciens, a novel chromate tolerant bacterium isolated from chromite mine soil. Chemosphere 96: 112–121. doi: 10.1016/j.chemosphere.2013.08.080
|
| [27] |
Javaid M, Sultan S (2013) Plant growth promotion traits and Cr (VI) reduction potentials of Cr (VI) resistant Streptomyces strains. J Basic Microbiol 53: 420–428. doi: 10.1002/jobm.201200032
|
| [28] | Bhattacharya A, Gupta A (2013) Evaluation of Acinetobacter sp. B9 for Cr(VI) resistance and detoxification with potential application in bioremediation of heavy-metals-rich industrial wastewater. Environ Sci Pollut Res 20: 6628–6637. |
| [29] |
Basu A, Ghosh SK, Saha R (2011) Effect of Some Non Functional Surfactants and Electrolytes on the Hexavalent Chromium Reduction by Glycerol: A Mechanistic Study. Tenside Surfactants Detergents 48: 453–458. doi: 10.3139/113.110152
|
| [30] |
Sultan S, Hasnain S (2006) Characterization of Ochrobactrum intermedium strain STCr-5 manifesting high level Cr(VI) resistance and reduction potential. Enzyme Microbial Technol 39: 883–888. doi: 10.1016/j.enzmictec.2006.01.019
|
| [31] |
Maret W (2013) Inhibitory zinc sites in enzymes. Biometals 26: 197–204. doi: 10.1007/s10534-013-9613-7
|
| [32] |
Opperman DJ, Heerden EV (2007) Aerobic Cr(VI) reduction by Thermus scotoductus strain SA-01. J Appl Microbiol 103: 1907–1913. doi: 10.1111/j.1365-2672.2007.03429.x
|
| [33] |
Wani R, Kodam KM, Gawai KR, et al. (2007) Chromate reduction by Burkholderia cepacia MCMB-821 isolated from the pristine habitat of alkaline crater lake. Appl Microbiol Biotechnol 75: 627–632. doi: 10.1007/s00253-007-0862-7
|
| [34] | Alam MZ, Ahmad S (2011) Toxic chromate reduction by resistant and sensitive bacteria isolated from tannery effluent contaminated soil. Ann Microbiol 43: 21–27. |
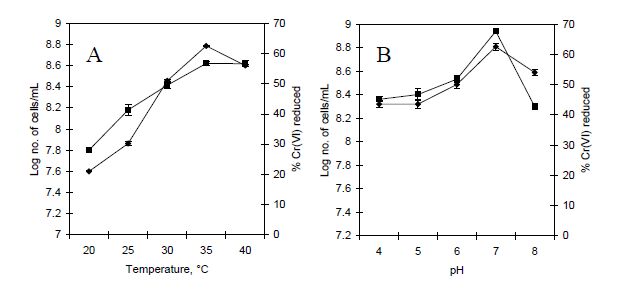
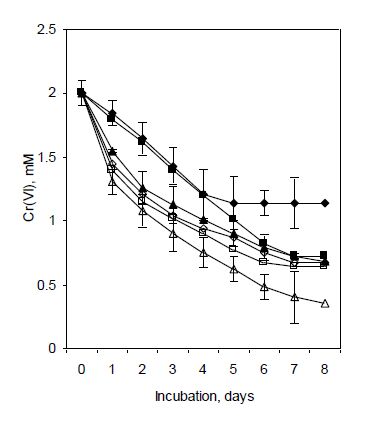
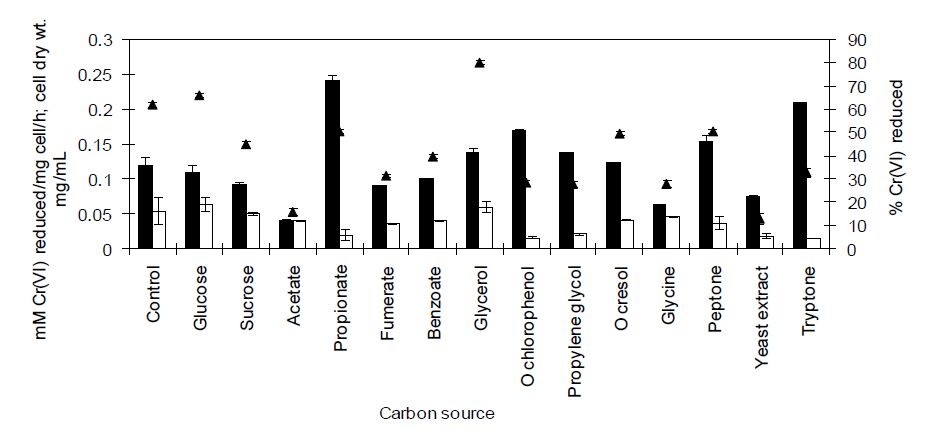
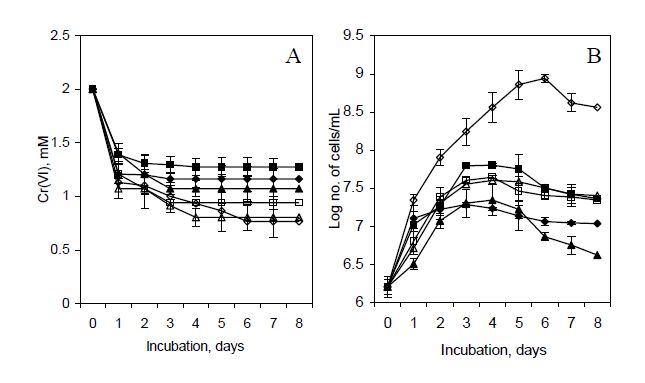
Figures(8) / Tables(2)

Satarupa Dey, Amal Kanti Paul. Assessment of heavy metal tolerance and hexavalent chromium reducing potential of Corynebacterium paurometabolum SKPD 1204 isolated from chromite mine seepage[J]. AIMS Bioengineering, 2016, 3(3): 337-351. doi: 10.3934/bioeng.2016.3.337







 DownLoad:
DownLoad: