Citation: Clara Navarrete, José L. Martínez. Non-conventional yeasts as superior production platforms for sustainable fermentation based bio-manufacturing processes[J]. AIMS Bioengineering, 2020, 7(4): 289-305. doi: 10.3934/bioeng.2020024

| [1] |
Carlson R (2016) Estimating the biotech sector's contribution to the US economy. Nat Biotechnol 34: 247-255. doi: 10.1038/nbt.3491

|
| [2] |
Parapouli M, Vasileiadis A, Afendra AS, et al. (2020) Saccharomyces cerevisiae and its industrial applications. AIMS Microbiology 6: 1-31. doi: 10.3934/microbiol.2020001
|
| [3] |
Baeshen NA, Baeshen MN, Sheikh A, et al. (2014) Cell factories for insulin production. Microb Cell Fact 13: 1-9. doi: 10.1186/s12934-014-0141-0
|
| [4] |
Hu W, Li WJ, Yang HQ, et al. (2019) Current strategies and future prospects for enhancing microbial production of citric acid. Appl Microbiol Biotechnol 103: 201-209. doi: 10.1007/s00253-018-9491-6
|
| [5] |
Park HS, Jun SC, Han KH, et al. (2017) Diversity, application, and synthetic biology of industrially important Aspergillus fungi. Adv Appl Microbiol 100: 161-202. doi: 10.1016/bs.aambs.2017.03.001
|
| [6] |
Fang H, Li D, Kang J, et al. (2018) Metabolic engineering of Escherichia coli for de novo biosynthesis of vitamin B12. Nat Commun 9: 4917. doi: 10.1038/s41467-018-07412-6
|
| [7] |
Zhao C, Zhang Y, Li Y (2019) Production of fuels and chemicals from renewable resources using engineered Escherichia coli. Biotechnol Adv 37: 107402. doi: 10.1016/j.biotechadv.2019.06.001
|
| [8] |
Hong KK, Nielsen J (2012) Metabolic engineering of Saccharomyces cerevisiae: a key cell factory platform for future biorefineries. Cell Mol Life Sci 69: 2671-2690. doi: 10.1007/s00018-012-0945-1
|
| [9] |
Pontrelli S, Chiu TYC, Lan EI, et al. (2018) Escherichia coli as a host for metabolic engineering. Metab Eng 50: 16-46. doi: 10.1016/j.ymben.2018.04.008
|
| [10] |
Thorwall S, Schwartz C, Chartron JW, et al. (2020) Stress-tolerant non-conventional microbes enable next-generation chemical biosynthesis. Nat Chem Biol 16: 113-121. doi: 10.1038/s41589-019-0452-x
|
| [11] | Rebello S, Abraham A, Madhavan A, et al. (2018) Non-conventional yeast cell factories for sustainable bioprocesses. FEMS Microbiol Lett 365: fny222. |
| [12] |
Löbs AK, Schwartz C, Wheeldon I (2017) Genome and metabolic engineering in non-conventional yeasts: current advances and applications. Syn Syst Biotechnol 2: 198-207. doi: 10.1016/j.synbio.2017.08.002
|
| [13] |
Jensen NB, Strucko T, Kildegaard KR, et al. (2014) EasyClone: method for iterative chromosomal integration of multiple genes in Saccharomyces cerevisiae. FEMS Yeast Res 14: 238-248. doi: 10.1111/1567-1364.12118
|
| [14] |
Vernis L, Poljak L, Chasles M, et al. (2001) Only centromeres can supply the partition system required for ARS function in the yeast Yarrowia lipolytica. J Mol Biol 305: 203-217. doi: 10.1006/jmbi.2000.4300
|
| [15] |
Lieber MR (2010) The mechanism of double-strand DNA break repair by the nonhomologous DNA end-joining pathway. Annu Rev Biochem 79: 181-211. doi: 10.1146/annurev.biochem.052308.093131
|
| [16] |
Flagfeldt DB, Siewers V, Huang L, et al. (2009) Characterization of chromosomal integration sites for heterologous gene expression in Saccharomyces cerevisiae. Yeast 26: 545-551. doi: 10.1002/yea.1705
|
| [17] |
Schwartz C, Shabbir-Hussain M, Frogue K, et al. (2017) Standardized markerless gene integration for pathway engineering in Yarrowia lipolytica. ACS Synth Biol 6: 402-409. doi: 10.1021/acssynbio.6b00285
|
| [18] |
Jinek M, Chylinski K, Fonfara I, et al. (2012) A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 337: 816-821. doi: 10.1126/science.1225829
|
| [19] |
Liu Z, Liang Y, Ang EL, et al. (2017) A new era of genome integration-simply cut and paste!. ACS Synth Biol 6: 601-609. doi: 10.1021/acssynbio.6b00331
|
| [20] |
Botstein D, Chervitz SA, Cherry JM (1997) Yeast as a model organism. Science 277: 1259-1260. doi: 10.1126/science.277.5330.1259
|
| [21] |
Liu HH, Ji XJ, Huang H (2015) Biotechnological applications of Yarrowia lipolytica: Past, present and future. Biotechnol Adv 33: 1522-1546. doi: 10.1016/j.biotechadv.2015.07.010
|
| [22] |
Xie D (2017) Integrating cellular and bioprocess engineering in the non-conventional yeast Yarrowia lipolytica for biodiesel production: A review. Front Bioeng Biotechnol 5: 65. doi: 10.3389/fbioe.2017.00065
|
| [23] |
Ledesma-Amaro R, Nicaud JM (2016) Metabolic engineering for expanding the substrate range of Yarrowia lipolytica. Trends Biotechnol 34: 798-809. doi: 10.1016/j.tibtech.2016.04.010
|
| [24] |
Jun S, XiaoFeng J, Yuan Z, et al. (2018) Expression, purification, crystallization, and diffraction analysis of a selenomethionyl lipase Lip8 from Yarrowia lipolytica. Prep Biochem Biotechnol 48: 213-217. doi: 10.1080/10826068.2016.1188316
|
| [25] |
Carly F, Steels S, Telek S, et al. (2018) Identification and characterization of EYD1, encoding an erythritol dehydrogenase in Yarrowia lipolytica and its application to bioconvert erythritol into erythrulose. Bioresour Technol 247: 963-969. doi: 10.1016/j.biortech.2017.09.168
|
| [26] |
Qiao K, Wasylenko TM, Zhou K, et al. (2017) Lipid production in Yarrowia lipolytica is maximized by engineering cytosolic redox metabolism. Nat Biotechnol 35: 173-177. doi: 10.1038/nbt.3763
|
| [27] |
Dujon B, Sherman D, Fischer G, et al. (2004) Genome evolution in yeasts. Nature 430: 35-44. doi: 10.1038/nature02579
|
| [28] |
Fickers P, Le Dall MT, Gaillardin C, et al. (2003) New disruption cassettes for rapid gene disruption and marker rescue in the yeast Yarrowia lipolytica. J Microbiol Methods 55: 727-737. doi: 10.1016/j.mimet.2003.07.003
|
| [29] |
Verbeke J, Beopoulos A, Nicaud JM (2013) Efficient homologous recombination with short length flanking fragments in Ku70 deficient Yarrowia lipolytica strains. Biotechnol Lett 35: 571-576. doi: 10.1007/s10529-012-1107-0
|
| [30] |
Wang HJ, Le Dall MT, Wach Y, et al. (1999) Evaluation of acylcoenzyme A oxidase (Aox) isozyme function in the n-alkane- assimilating yeast Yarrowia lipolytica. J Bacteriol 181: 5140-5148. doi: 10.1128/JB.181.17.5140-5148.1999
|
| [31] |
Gao S, Tong Y, Wen Z, et al. (2016) Multiplex gene editing of the Yarrowia lipolytica genome using the CRISPR-Cas9 system. J Ind Microbiol Biotechnol 43: 1085-1093. doi: 10.1007/s10295-016-1789-8
|
| [32] |
Schwartz CM, Shabbir-Hussain M, Blenner M, et al. (2016) Synthetic RNA polymerase III promoters facilitate high-efficiency CRISPR-Cas9-mediated genome editing in Yarrowia lipolytica. ACS Synth Biol 5: 356-359. doi: 10.1021/acssynbio.5b00162
|
| [33] |
Yaguchi A, Rives D, Blenner M (2017) New kids on the block: emerging oleaginous yeast of biotechnological importance. AIMS Microbiology 3: 227-247. doi: 10.3934/microbiol.2017.2.227
|
| [34] |
Moon NJ, Hammond EG, Glatz BA (1978) Conversion of cheese whey and whey permeate to oil and single-cell protein. J Dairy Sci 61: 1537-1547. doi: 10.3168/jds.S0022-0302(78)83762-X
|
| [35] |
Bednarski W, Leman J, Tomasik J (1986) Utilization of beet molasses and whey for fat biosynthesis by a yeast. Agr Wastes 18: 19-26. doi: 10.1016/0141-4607(86)90104-6
|
| [36] |
Christophe G, Deo JL, Kumar V, et al. (2012) Production of oils from acetic acid by the oleaginous yeast Cryptococcus curvatus. Appl Biochem Biotechnol 167: 1270-1279. doi: 10.1007/s12010-011-9507-5
|
| [37] |
Yu X, Zheng Y, Dorgan KM, et al. (2011) Oil production by oleaginous yeasts using the hydrolysate from pretreatment of wheat straw with dilute sulfuric acid. Bioresour Technol 102: 6134-6140. doi: 10.1016/j.biortech.2011.02.081
|
| [38] |
Zheng Y, Chi Z, Ahring BK, et al. (2012) Oleaginous yeast Cryptococcus curvatus for biofuel production: Ammonia's effect. Biomass Bioenergy 37: 114-121. doi: 10.1016/j.biombioe.2011.12.022
|
| [39] |
Zhanga X, Chena J, Wu D, et al. (2019) Economical lipid production from Trichosporon oleaginosus via dissolved oxygen adjustment and crude glycerol addition. Bioresour Technol 273: 288-296. doi: 10.1016/j.biortech.2018.11.033
|
| [40] |
Chen J, Zhang X, Drogui P, et al. (2018) The pH-based fed-batch for lipid production from Trichosporon oleaginosus with crude glycerol. Bioresour Technol 259: 237-243. doi: 10.1016/j.biortech.2018.03.045
|
| [41] |
Yaguchi A, Robinson A, Mihealsick E, et al. (2017) Metabolism of aromatics by Trichosporon oleaginosus while remaining oleaginous. Microb Cell Fact 16: 206. doi: 10.1186/s12934-017-0820-8
|
| [42] |
Kourist R, Bracharz F, Lorenzen J, et al. (2015) Genomics and transcriptomics analyses of the oil-accumulating basidiomycete yeast Trichosporon oleaginosus: insights into substrate utilization and alternative evolutionary trajectories of fungal mating systems. MBio 6: e0091815. doi: 10.1128/mBio.00918-15
|
| [43] |
Görner C, Redai V, Bracharz F, et al. (2016) Genetic engineering and production of modified fatty acids by the non-conventional oleaginous yeast Trichosporon oleaginosus ATCC 20509. Green Chem 18: 2037-2046. doi: 10.1039/C5GC01767J
|
| [44] |
Nurcholis M, Lertwattanasakul N, Rodrussamee N, et al. (2020) Integration of comprehensive data and biotechnological tools for industrial applications of Kluyveromyces marxianus. Appl Microbiol Biotechnol 104: 475-488. doi: 10.1007/s00253-019-10224-3
|
| [45] |
Radecka D, Mukherjee V, Mateo RQ, et al. (2015) Looking beyond Saccharomyces: the potential of non-conventional yeast species for desirable traits in bioethanol fermentation. FEMS Yeast Res 15: fov053. doi: 10.1093/femsyr/fov053
|
| [46] |
Nonklang S, Abdel-Banat BMA, Cha-aim K, et al. (2008) High-temperature ethanol fermentation and transformation with linear DNA in the thermotolerant yeast Kluyveromyces marxianus DMKU3–1042. Appl Environ Microb 74: 7514-7521. doi: 10.1128/AEM.01854-08
|
| [47] |
Rouwenhorst RJ, Visser LE, Baan AA, et al. (1988) Production, distribution, and kinetic properties of inulinase in continuous cultures of Kluyveromyces marxianus CBS 6556. Appl Environ Microb 54: 1131-1137. doi: 10.1128/AEM.54.5.1131-1137.1988
|
| [48] |
Martins DBG, de Souza CG, Simões DA, et al. (2002) The betagalactosidase activity in Kluyveromyces marxianus CBS6556 decreases by high concentrations of galactose. Curr Microbiol 44: 379-382. doi: 10.1007/s00284-001-0052-2
|
| [49] | Kosaka T, Lertwattanasakul N, Rodrussamee N, et al. (2018) Potential of thermotolerant ethanologenic yeasts isolated from ASEAN countries and their application in high-temperature fermentation. Fuel Ethanol Production from Sugarcane London: IntechOpen, 121-154. |
| [50] |
Fonseca GG, Heinzle E, Wittmann C, et al. (2008) The yeast Kluyveromyces marxianus and its biotechnological potential. Appl Microbiol Biotechnol 79: 339-354. doi: 10.1007/s00253-008-1458-6
|
| [51] |
Kim TY, Lee SW, Oh MK (2014) Biosynthesis of 2-phenylethanol from glucose with genetically engineered Kluyveromyces marxianus. Enzym Microb Technol 61: 44-47. doi: 10.1016/j.enzmictec.2014.04.011
|
| [52] |
Jeong H, Lee D-H, Kim SH, et al. (2012) Genome sequence of the thermotolerant yeast Kluyveromyces marxianus var marxianus KCTC 17555. Eukaryot Cell 11: 1584-1585. doi: 10.1128/EC.00260-12
|
| [53] |
Heo P, Yang TJ, Chung SC, et al. (2013) Simultaneous integration of multiple genes into the Kluyveromyces marxianus chromosome. J Biotechnol 167: 323-325. doi: 10.1016/j.jbiotec.2013.06.020
|
| [54] |
Lertwattanasakul N, Kosaka T, Hosoyama A, et al. (2015) Genetic basis of the highly efficient yeast Kluyveromyces marxianus: complete genome sequence and transcriptome analyses. Biotechnol Biofuels 8: 47. doi: 10.1186/s13068-015-0227-x
|
| [55] |
Schabort DTWP, Letebele PK, Steyn L, et al. (2016) Differential RNA-seq, multi-network analysis and metabolic regulation analysis of Kluyveromyces marxianus reveals a compartmentalised response to xylose. PLoS One 11: e0156242. doi: 10.1371/journal.pone.0156242
|
| [56] |
Rollero S, Bloem A, Ortiz-Julien A, et al. (2019) A comparison of the nitrogen metabolic networks of Kluyveromyces marxianus and Saccharomyces cerevisiae. Environ Microbiol 21: 4076-4091. doi: 10.1111/1462-2920.14756
|
| [57] |
Pentjuss A, Stalidzans E, Liepins J, et al. (2017) Model-based biotechnological potential analysis of Kluyveromyces marxianus central metabolism. J Ind Microbiol Biotechnol 44: 1177-1190. doi: 10.1007/s10295-017-1946-8
|
| [58] |
Marcišauskas S, Ji B, Nielsen J (2019) Reconstruction and analysis of a Kluyveromyces marxianus genome-scale metabolic model. BMC Bioinformatics 20: 551. doi: 10.1186/s12859-019-3134-5
|
| [59] |
Sakihama Y, Hidese R, Hasunuma T, et al. (2019) Increased flux in acetyl-CoA synthetic pathway and TCA cycle of Kluyveromyces marxianus under respiratory conditions. Sci Rep 9: 5319. doi: 10.1038/s41598-019-41863-1
|
| [60] | Nambu-Nishida Y, Nishida K, Hasunuma T, et al. (2017) Development of a comprehensive set of tools for genome engineering in a cold- and thermo-tolerant Kluyveromyces marxianus yeast strain. SciRep 7: 8993. |
| [61] |
Löbs AK, Schwartz C, Thorwal S, et al. (2018) Highly multiplexed CRISPRi repression of respiratory functions enhances mitochondrial localized ethyl acetate biosynthesis in Kluyveromyces marxianus. ACS Synth Biol 7: 2647-2655. doi: 10.1021/acssynbio.8b00331
|
| [62] |
Rajkumar AS, Varela JA, Juergens H, et al. (2019) Biological parts for Kluyveromyces marxianus synthetic biology. Front Bioeng Biotechnol 7: 97. doi: 10.3389/fbioe.2019.00097
|
| [63] |
Echeverrigaray S, Randon M, daSilva K, et al. (2013) Identification and characterization of non-saccharomyces spoilage yeasts isolated from Brazilian wines. World J Microb Biot 29: 1019-1027. doi: 10.1007/s11274-013-1265-9
|
| [64] |
Rozpedowska E, Hellborg L, Ishchuk OP, et al. (2011) Parallel evolution of the make-accumulate-consume strategy in Saccharomyces and Dekkera yeasts. Nat Commun 2: 302. doi: 10.1038/ncomms1305
|
| [65] |
Piskur J, Ling Z, Marcet-Houben M, et al. (2012) The genome of wine yeast Dekkera bruxellensis provides a tool to explore its food-related properties. Int J Food Microbiol 157: 202-209. doi: 10.1016/j.ijfoodmicro.2012.05.008
|
| [66] | Conterno L, Joseph CML, Arvik TJ, et al. (2006) Genetic and physiological characterization of Brettanomyces bruxellensis strains isolated from wines. Am J Enol Viticult 57: 139-147. |
| [67] |
Wijsman MR, van Dijken JP, van Kleeff BHA, et al. (1984) Inhibition of fermentation and growth in batch cultures of the yeast Brettanomyces intermedius upon a shift from aerobic to anaerobic conditions (Custers effect). Anton Leeuw 50: 183-192. doi: 10.1007/BF00400180
|
| [68] |
Brandam C, Castro-Martínez C, Délia ML, et al. (2008) Effect of temperature on Brettanomyces bruxellensis: metabolic and kinetic aspects. Can J Microbiol 54: 11-18. doi: 10.1139/W07-126
|
| [69] |
Taillandier P, Lai QP, Julien-Ortiz A, et al. (2014) Interactions between Torulaspora delbrueckii and Saccharomyces cerevisiae in wine fermentation: influence of inoculation and nitrogen content. World J Microb Biot 30: 1959-1967. doi: 10.1007/s11274-014-1618-z
|
| [70] |
Blomqvist J, Passoth V (2015) Dekkera bruxellensis--spoilage yeast with biotechnological potential, and a model for yeast evolution, physiology and competitiveness. FEMS Yeast Res 15: fov021. doi: 10.1093/femsyr/fov021
|
| [71] |
Woolfit M, Rozpedowska E, Piskur J, et al. (2007) Genome survey sequencing of the wine spoilage yeast Dekkera (Brettanomyces) bruxellensis. Eukaryot Cell 6: 721-733. doi: 10.1128/EC.00338-06
|
| [72] |
Curtin CD, Borneman AR, Chambers PJ, et al. (2012) Denovo assembly and analysis of the heterozygous triploid genome of the wine spoilage yeast Dekkera bruxellensis AWRI1499. PLoS One 7: e33840. doi: 10.1371/journal.pone.0033840
|
| [73] |
Miklenic M, Stafa A, Bajic A, et al. (2013) Genetic transformation of the yeast Dekkera/Brettanomyces bruxellensis with nonhomologous DNA. J Microbiol Biotechnol 23: 674-680. doi: 10.4014/jmb.1211.11047
|
| [74] |
Tiukova IA, Petterson ME, Tellgren-Roth C, et al. (2013) Transcriptome of the alternative ethanol production strain Dekkera bruxellensis CBS 11270 in sugar limited, low oxygen cultivation. PLoS One 8: e58455. doi: 10.1371/journal.pone.0058455
|
| [75] |
Oberoi HS, Babbar N, Sandhu SK, et al. (2012) Ethanol production from alkali-treated rice straw via simultaneous saccharification and fermentation using newly isolated thermotolerant Pichia kudriavzevii HOP-1. J Ind Microbiol Biot 39: 557-566. doi: 10.1007/s10295-011-1060-2
|
| [76] |
Schnierda T, Bauer FF, Divol B, et al. (2014) Optimization of carbon and nitrogen medium components for biomass production using non-Saccharomyces wine yeasts. Lett Appl Microbiol 58: 478-485. doi: 10.1111/lam.12217
|
| [77] |
Daniel HM, Vrancken G, Takrama JF, et al. (2009) Yeast diversity of Ghanaian cocoa bean heap fermentations. FEMS Yeast Res 9: 774-783. doi: 10.1111/j.1567-1364.2009.00520.x
|
| [78] |
Kitagawa T, Tokuhiro K (2010) Construction of a β-glucosidase expression system using the multistress-tolerant yeast Issatchenkia orientalis. Appl Microbiol Biot 87: 1841-1853. doi: 10.1007/s00253-010-2629-9
|
| [79] |
Kwon YJ, Ma A-Z, Li Q, et al. (2011) Effect of lignocellulosic inhibitory compounds on growth and ethanol fermentation of newly-isolated thermotolerant Issatchenkia orientalis. Bioresour Technol 102: 8099-8104. doi: 10.1016/j.biortech.2011.06.035
|
| [80] |
Dandi ND, Dandi BN, Chaudhari AB (2013) Bioprospecting of thermos and osmo-tolerant fungi from mango pulp-peel compost for bioethanol production. Anton Leeuw 103: 723-736. doi: 10.1007/s10482-012-9854-4
|
| [81] |
Chan GF, Gan HM, Ling HL, et al. (2012) Genome sequence of Pichia kudriavzevii M12, a potential producer of bioethanol and phytase. Eukaryot Cell. 11: 1300-1301. doi: 10.1128/EC.00229-12
|
| [82] | Finley KR, Huryta JM, Mastel BM, et al. (2013) Compositions and methods for succinate production. US patent 2013. |
| [83] | Rush BJ, Fosmer AM (2014) Methods for succinate production. US patent 2014. |
| [84] |
Prista C, Michán C, Miranda IM, et al. (2016) The halotolerant Debaryomyces hansenii, the Cinderella of non-conventional yeasts. Yeast 33: 523-533. doi: 10.1002/yea.3177
|
| [85] |
Almagro A, Prista C, Castro S, et al. (2000) Effects of salt on Debaryomyces hansenii and Saccharomyces cerevisiae under stress conditions. Int J Food Microbiol 56: 191-197. doi: 10.1016/S0168-1605(00)00220-8
|
| [86] |
Papouskova K, Sychrova H (2007) The co-action of osmotic and high temperature stresses results in a growth improvement of Debaryomyces hansenii cells. Int J Food Microbiol 118: 1-7. doi: 10.1016/j.ijfoodmicro.2007.04.005
|
| [87] |
Navarrete C, Siles A, Martínez JL, et al. (2009) Oxidative stress sensitivity in Debaryomyces hansenii. FEMS Yeast Res 9: 582-590. doi: 10.1111/j.1567-1364.2009.00500.x
|
| [88] |
Gustafsson L, Norkrans B (1976) On the mechanism of salt tolerance: production of glycerol and heat during growth of Debaryomyces hansenii. Arch Microbiol 110: 177-183. doi: 10.1007/BF00690226
|
| [89] |
Adler L, Blomberg A, Nilsson A (1985) Glycerol metabolism and osmoregulation in the salt-tolerant yeast Debaryomyces hansenii. J Bacteriol 162: 300-306. doi: 10.1128/JB.162.1.300-306.1985
|
| [90] |
Breuer U, Harms H (2006) Debaryomyces hansenii—an extremophilic yeast with biotechnological potential. Yeast 23: 415-437. doi: 10.1002/yea.1374
|
| [91] |
Ramfrez-Orozco M, Hernandez-Saavedra N, Ochoa JL (2001) Debaryomyces hansenii growth in nonsterile seawater ClO2-peptone-containing medium. Can J Microbiol 47: 676-679. doi: 10.1139/w01-056
|
| [92] |
Ricaurte ML, Govind NS (1999) Construction of plasmid vectors and transformation of the marine yeast Debaryomyces hansenii. Mar Biotechnol 1: 15-19. doi: 10.1007/PL00011745
|
| [93] | Voronovsky A, Abbas C, Fayura L, et al. (2002) Development of a transformation system for the flavinogenic yeast. FEMS Yeast Res 2: 381-388. |
| [94] |
Terentiev Y, Pico AH, Boer E, et al. (2004) A wide-range integrative yeast expression vector system based on Arxula adeninivorans-derived elements. J Ind Microbiol Biotechnol 31: 223-228. doi: 10.1007/s10295-004-0142-9
|
| [95] |
Dmytruk KV, Voronovsky AY, Sibirny AA (2006) Insertion mutagenesis of the yeast Candida famata (Debaryomyces hansenii) by random integration of linear DNA fragments. Curr Genet 50: 183-191. doi: 10.1007/s00294-006-0083-0
|
| [96] |
Minhas A, Biswas D, Mondal AK (2009) Development of host and vector for high-efficiency transformation and gene disruption in Debaryomyces hansenii. FEMS Yeast Res 9: 95-102. doi: 10.1111/j.1567-1364.2008.00457.x
|
| [97] | Rhee SJ, Lee CYJ, Kim KK, et al. (2003) Comparison of the traditional (Samhaeju) and industrial (Chongju) rice wine brewing in Korea. Food Sci Biotechnol 12: 242-247. |
| [98] | Sahm H (1977) Metabolism of methanol by yeast. Adv Biochem Eng 6: 77-103. |
| [99] |
Hansen H, Hollenberg CP (1996) Hansenula polymorpha (Pichia angusta). Nonconventional Yeasts in Biotechnology Heidelberg: Springer, 293-311. doi: 10.1007/978-3-642-79856-6_9
|
| [100] |
Voronovsky AY, Rohulya OV, Abbas CA, et al. (2009) Development of strains of the thermotolerant yeast Hansenula polymorpha capable of alcoholic fermentation of starch and xylan. Metab Eng 11: 234-242. doi: 10.1016/j.ymben.2009.04.001
|
| [101] |
Ryabova OB, Chmil OM, Sibirny AA (2003) Xylose and cellobiose fermentation to ethanol by the thermotolerant methylotrophic yeast Hansenula polymorpha. FEMS Yeast Res 4: 157-164. doi: 10.1016/S1567-1356(03)00146-6
|
| [102] |
Dmytruk K, Kurylenko O, Ruchala J, et al. (2017) Development of the thermotolerant methylotrophic yeast Hansenula polymorpha as efficient ethanol producer. Yeast Diversity in Human Welfare Singapore: Springer, 257-282. doi: 10.1007/978-981-10-2621-8_11
|
| [103] | van der Klei IJ, Yurimoto H, Sakai Y, et al. (2006) The significance of peroxisomes in methanol metabolism in methylotrophic yeast. Bba-Mol Cell Res 1763: 1453-1462. |
| [104] |
Siverio JM (2002) Assimilation of nitrate by yeasts. Fems Microbiol Rev 26: 277-284. doi: 10.1111/j.1574-6976.2002.tb00615.x
|
| [105] |
Gidijala L, Kiel JA, Douma RD, et al. (2009) An engineered yeast efficiently secreting penicillin. PLoS One 4: e8317. doi: 10.1371/journal.pone.0008317
|
| [106] |
Kunze G, Kang HA, Gellissen G, et al. (2009) Hansenula polymorpha (Pichia angusta): biology and applications. Yeast Biotechnology: Diversity and Applications Dordrecht: Springer, 47-64. doi: 10.1007/978-1-4020-8292-4_3
|
| [107] | Brierley RA, Davis GR, Holtz GC, et al. (1997) Production of insulin-like growth factor-1 in methylotrophic yeast cells. US patent 1997. |
| [108] |
Janowicz ZA, Melber K, Merckelbach A, et al. (1991) Simultaneous expression of the S and L surface antigens of hepatitis B, and formation of mixed particles in the methylotrophic yeast, Hansenula polymorpha. Yeast 7: 431-443. doi: 10.1002/yea.320070502
|
| [109] |
Kulkarni PS, Raut SK, Patki PS, et al. (2006) Immunogenicity of a new, low-cost recombinant hepatitis B vaccine derived from Hansenula polymorpha in adults. Vaccine 24: 3457-3460. doi: 10.1016/j.vaccine.2006.02.008
|
| [110] |
Ramenazi-Rad M, Hollenberg CP, Lauber J, et al. (2003) The Hansenula polymorpha (strain CBS4732) genome sequencing and analysis. FEMS Yeast Res 4: 207-215. doi: 10.1016/S1567-1356(03)00125-9
|
| [111] |
Krasovska O, Stasyk OG, Nahorny VO, et al. (2007) Glucose-induced production of recombinant proteins in Hansenula polymorpha mutants deficient in catabolite repression. Biotechnol Bioeng 97: 858-870. doi: 10.1002/bit.21284
|
| [112] |
Saraya R, Krikken AM, Kiel JAKW, et al. (2012) Novel genetic tools for Hansenula polymorpha. FEMS Yeast Res 12: 271-278. doi: 10.1111/j.1567-1364.2011.00772.x
|
| [113] |
Heo JH, Hong WK, Cho EY, et al. (2003) Properties of the Hansenula polymorpha-derived constitutive GAP promoter, assessed using an HAS reporter gene. FEMS Yeast Res 4: 175-184. doi: 10.1016/S1567-1356(03)00150-8
|
| [114] |
Bogdanova AI, Agaphonov MO, Ter-Avanesyan MD (1995) Plasmid reorganization during integrative transformation in Hansenula polymorpha. Yeast 11: 343-353. doi: 10.1002/yea.320110407
|
| [115] |
Numamoto M, Maekawa H, Kaneko Y (2017) Efficient genome editing by CRISPR/Cas9 with a tRNA-sgRNA fusion in the methylotrophic yeast Ogataea polymorpha. J Biosci Bioeng 124: 487-492. doi: 10.1016/j.jbiosc.2017.06.001
|
| [116] |
Blazeck J, Hill A, Liu L, et al. (2014) Harnessing Yarrowia lipolytica lipogenesis to create a platform for lipid and biofuel production. Nat Commun 5: 3131. doi: 10.1038/ncomms4131
|
| [117] |
Tai M, Stephanopoulos G (2013) Engineering the push and pull of lipid biosynthesis in oleaginous yeast Yarrowia lipolytica for biofuel production. Metab Eng 15: 1-9. doi: 10.1016/j.ymben.2012.08.007
|
| [118] |
Förster A, Aurich A, Mauersberger S, et al. (2007) Citric acid production from sucrose using a recombinant strain of the yeast Yarrowia lipolytica. Appl Microbiol Biotechnol 75: 1409-1417. doi: 10.1007/s00253-007-0958-0
|
| [119] |
Mironczuk AM, Furgala J, Rakicka M, et al. (2014) Enhanced production of erythritol by Yarrowia lipolytica on glycerol in repeated batch cultures. J Ind Microbiol Biotechnol 41: 57-64. doi: 10.1007/s10295-013-1380-5
|
| [120] |
Yovkova V, Otto C, Aurich A, et al. (2014) Engineering the α-ketoglutarate overproduction from raw glycerol by overexpression of the genes encoding NADP+-dependent isocitrate dehydrogenase and pyruvate carboxylase in Yarrowia lipolytica. Appl Microbiol Biotechnol 98: 2003-2013. doi: 10.1007/s00253-013-5369-9
|
| [121] |
Matthäus F, Ketelhot M, Gatter M, et al. (2014) Production of lycopene in the non-carotenoid-producing yeast Yarrowia lipolytica. Appl Environ Microbiol 80: 1660-1669. doi: 10.1128/AEM.03167-13
|
| [122] |
Xue Z, Sharpe PL, Hong SP, et al. (2013) Production of omega-3 eicosapentaenoic acid by metabolic engineering of Yarrowia lipolytica. Nat Biotechnol 31: 734-740. doi: 10.1038/nbt.2622
|
| [123] |
Cheon Y, Kim JS, Park JB, et al. (2014) A biosynthetic pathway for hexanoic acid production in Kluyveromyces marxianus. J Biotechnol 182: 30-36. doi: 10.1016/j.jbiotec.2014.04.010
|
| [124] |
Ramirez-Zavala B, Mercado-Flores Y, Hernandez-Rodriguez C, et al. (2004) Purification and characterization of a lysine aminopeptidase from Kluyveromyces marxianus. FEMS Microbiol Lett 235: 369-375. doi: 10.1111/j.1574-6968.2004.tb09612.x
|
| [125] |
Ramirez-Zavala B, Mercado-Flores Y, Hernandez-Rodriguez C, et al. (2004) Purification and characterization of a serine carboxypeptidase from Kluyveromyces marxianus. Int J Food Microbiol 91: 245-252. doi: 10.1016/S0168-1605(03)00409-4
|
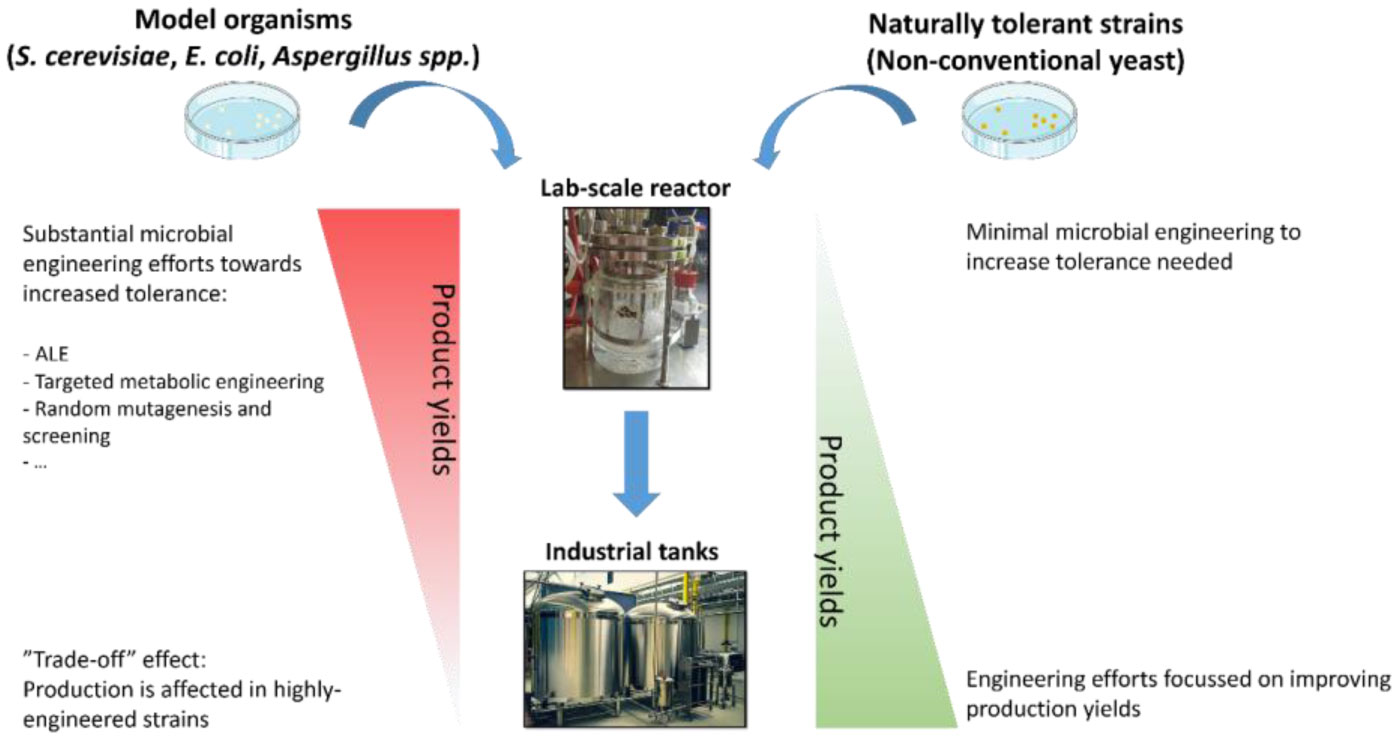
Figures(1) / Tables(1)

Clara Navarrete, José L. Martínez. Non-conventional yeasts as superior production platforms for sustainable fermentation based bio-manufacturing processes[J]. AIMS Bioengineering, 2020, 7(4): 289-305. doi: 10.3934/bioeng.2020024







 DownLoad:
DownLoad: