Citation: Mingyong Xiong, Ping Yu, Jingyu Wang, Kechun Zhang. Improving Engineered Escherichia coli strains for High-level Biosynthesis of Isobutyrate[J]. AIMS Bioengineering, 2015, 2(2): 60-74. doi: 10.3934/bioeng.2015.2.60

| [1] | McFarlane J, Robinson S (2007) Survey of Alternative Feedstocks for Commodity Chemical Manufacturing. Available from: http://infoornlgov/sites/publications/files/Pub8760pdf. |
| [2] |
Atsumi S, Hanai T, Liao JC (2008) Non-fermentative pathways for synthesis of branched-chain higher alcohols as biofuels. Nature 451: 86-89. doi: 10.1038/nature06450

|
| [3] |
Causey TB, Zhou S, Shanmugam KT, et al. (2003) Engineering the metabolism of Escherichia coli W3110 for the conversion of sugar to redox-neutral and oxidized products: homoacetate production. Proc Natl Acad Sci USA 100: 825-832. doi: 10.1073/pnas.0337684100
|
| [4] | Pfeifer BA, Admiraal SJ, Gramajo H, et al. (2001) Biosynthesis of complex polyketides in a metabolically engineered strain of E. coli. Science 291: 1790-1792. |
| [5] |
Steen EJ, Kang Y, Bokinsky G, et al. (2010) Microbial production of fatty-acid-derived fuels and chemicals from plant biomass. Nature 463: 559-562. doi: 10.1038/nature08721
|
| [6] |
Yan Y, Chemler J, Huang L, et al. (2005) Metabolic engineering of anthocyanin biosynthesis in Escherichia coli. Appl Environ Microbiol 71: 3617-3623. doi: 10.1128/AEM.71.7.3617-3623.2005
|
| [7] |
Zha W, Shao Z, Frost JW, et al. (2004) Rational pathway engineering of type I fatty acid synthase allows the biosynthesis of triacetic acid lactone from D-glucose in vivo. J Am Chem Soc 126: 4534-4535. doi: 10.1021/ja0317271
|
| [8] |
Alper H, Miyaoku K, Stephanopoulos G (2005) Construction of lycopene-overproducing Escherichia coli strains by combining systematic and combinatorial gene knockout targets. Nat Biotechnol 23: 612-616. doi: 10.1038/nbt1083
|
| [9] |
Bastian S, Liu X, Meyerowitz JT, et al. (2011) Engineered ketol-acid reductoisomerase and alcohol dehydrogenase enable anaerobic 2-methylpropan-1-ol production at theoretical yield in Escherichia coli. Metab Eng 13: 345-352. doi: 10.1016/j.ymben.2011.02.004
|
| [10] |
Park JH, Lee KH, Kim TY, et al. (2007) Metabolic engineering of Escherichia coli for the production of L-valine based on transcriptome analysis and in silico gene knockout simulation. Proc Natl Acad Sci USA 104: 7797. doi: 10.1073/pnas.0702609104
|
| [11] |
Zhang K, Woodruff AP, Xiong M, et al. (2011) A synthetic metabolic pathway for production of the platform chemical isobutyric acid. ChemSusChem 4: 1068-1070. doi: 10.1002/cssc.201100045
|
| [12] | Godshall MA Value-Added Products for a Sustainable Sugar Industry. Sustainability of the Sugar and Sugar-Ethanol Industries: Eggleston, G.; ACS Symposium Series 1058; American Chemical Society: Washington, DC, 2010 1253-1268. |
| [13] | Screening Information Data Set (SIDS) for High Production Volume Chemicals, Organization for Economic Cooperation and Development. (2005) Available from: http://wwwinchemorg/documents/sids/sids/25265774pdf. |
| [14] |
Lee IY, Hong WK, Hwang YB, et al. (1996) Production of D-β-hydroxyisobutyric acid from isobutyric acid by Candida rugosa. J Ferment Bioeng 81: 79-82. doi: 10.1016/0922-338X(96)83126-6
|
| [15] |
Millet JMM (1998) FePO catalysts for the selective oxidative dehydrogenation of isobutyric acid into methacrylic acid. Catal Rev Sci Eng 40: 1-38. doi: 10.1080/01614949808007104
|
| [16] | Marx A, Poetter M, Buchholz S, et al. (2007) Microbiological Production of 3-Hydroxyisobutyric Acid. US Patent App 20: 773. |
| [17] |
Nagai K (2001) New developments in the production of methyl methacrylate. Appl Catal A-Gen 221: 367-377. doi: 10.1016/S0926-860X(01)00810-9
|
| [18] |
Atsumi S, Wu TY, Eckl EM, et al. (2010) Engineering the isobutanol biosynthetic pathway in Escherichia coli by comparison of three aldehyde reductase/alcohol dehydrogenase genes. Appl Microbiol Biotechnol 85: 651-657. doi: 10.1007/s00253-009-2085-6
|
| [19] | Baba T, Ara T, Hasegawa M, et al. (2006) Construction of Escherichia coli K-12 in-frame, single-gene knockout mutants: the Keio collection. Mol Syst Biol 2: 10.1038. |
| [20] |
Atsumi S, Wu TY, Eckl EM, et al. (2010) Engineering the isobutanol biosynthetic pathway in Escherichia coli by comparison of three aldehyde reductase/alcohol dehydrogenase genes. Appl Microbiol Biotechnol 85: 651-657. doi: 10.1007/s00253-009-2085-6
|
| [21] |
Atsumi S, Wu T-Y, Eckl E-M, et al. (2010) Engineering the isobutanol biosynthetic pathway in Escherichia coli by comparison of three aldehyde reductase/alcohol dehydrogenase genes. Appl Microbiol Biotechnol 85: 651-657. doi: 10.1007/s00253-009-2085-6
|
| [22] | Kallio P, Pásztor A, Thiel K, et al. (2014) An engineered pathway for the biosynthesis of renewable propane. Nat Commun 5. |
| [23] |
Rodriguez GM, Atsumi S (2012) Isobutyraldehyde production from Escherichia coli by removing aldehyde reductase activity. Microb Cell Fact 11: 90. doi: 10.1186/1475-2859-11-90
|
| [24] |
Åkesson M, Hagander P, Axelsson JP (2001) Avoiding acetate accumulation in Escherichia coli cultures using feedback control of glucose feeding. Biotechnol Bioeng 73: 223-230. doi: 10.1002/bit.1054
|
| [25] |
Thompson BG, Kole M, Gerson DF (1985) Control of ammonium concentration in Escherichia coli fermentations. Biotechnol Bioeng 27: 818-824. doi: 10.1002/bit.260270610
|
| [26] | Riesenberg D, Menzel K, Schulz V, et al. (1990) High cell density fermentation of recombinant Escherichia coli expressing human interferon alpha 1. Appl Environ Microbiol 34: 77-82. |
| [27] |
Eiteman MA, Altman E (2006) Overcoming acetate in Escherichia coli recombinant protein fermentations. Trends Biotechnol 24: 530-536. doi: 10.1016/j.tibtech.2006.09.001
|
| [28] |
Koh BT, Nakashimada U, Pfeiffer M, et al. (1992) Comparison of acetate inhibition on growth of host and recombinant E. coli K12 strains. Biotechnol Lett 14: 1115-1118. doi: 10.1007/BF01027012
|
| [29] |
Zhu Y, Eiteman M, DeWitt K, et al. (2007) Homolactate fermentation by metabolically engineered Escherichia coli strains. Appl Environ Microbiol 73: 456-464. doi: 10.1128/AEM.02022-06
|
| [30] | Stephanopoulos G, Aristidou AA, Nielsen JH, et al. (1998) Metabolic engineering: principles and methodologies: Academic Press. |
| [31] |
Warnecke T, Gill RT (2005) Organic acid toxicity, tolerance, and production in Escherichia coli biorefining applications. Microb Cell Fact 4: 25. doi: 10.1186/1475-2859-4-25
|
| [32] |
Yim H, Haselbeck R, Niu W, et al. (2011) Metabolic engineering of Escherichia coli for direct production of 1, 4-butanediol. Nat Chem Biol 7: 445-452. doi: 10.1038/nchembio.580
|
| [33] |
Qian ZG, Xia XX, Lee SY (2011) Metabolic engineering of Escherichia coli for the production of cadaverine: a five carbon diamine. Biotechnol Bioeng 108: 93-103. doi: 10.1002/bit.22918
|
| [34] |
Moon TS, Dueber JE, Shiue E, et al. (2010) Use of modular, synthetic scaffolds for improved production of glucaric acid in engineered E. coli. Metab Eng 12: 298-305. doi: 10.1016/j.ymben.2010.01.003
|
| [35] |
Dellomonaco C, Clomburg JM, Miller EN, et al. (2011) Engineered reversal of the β-oxidation cycle for the synthesis of fuels and chemicals. Nature 476: 355-359. doi: 10.1038/nature10333
|
| [36] |
McKenna R, Nielsen DR (2011) Styrene biosynthesis from glucose by engineered E. coli. Metab Eng 13: 544-554. doi: 10.1016/j.ymben.2011.06.005
|
| [37] |
Xiong M, Schneiderman DK, Bates FS, et al. (2014) Scalable production of mechanically tunable block polymers from sugar. Proc Natl Acad Sci U S A 111: 8357-8362. doi: 10.1073/pnas.1404596111
|
| [38] |
Datta R, Henry M (2006) Lactic acid: recent advances in products, processes and technologies a review. J Chem Technol Biotechnol 81: 1119-1129. doi: 10.1002/jctb.1486
|
| [39] |
Lin H, Bennett GN, San KY (2005) Metabolic engineering of aerobic succinate production systems in Escherichia coli to improve process productivity and achieve the maximum theoretical succinate yield. Metab Eng 7: 116-127. doi: 10.1016/j.ymben.2004.10.003
|
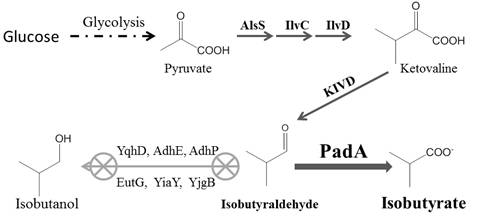
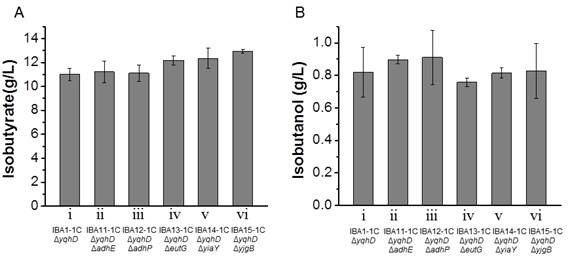
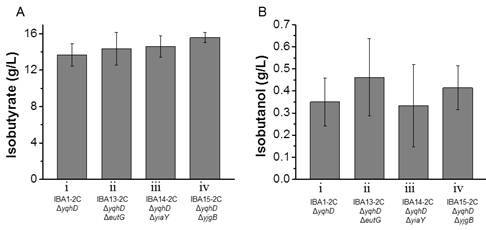
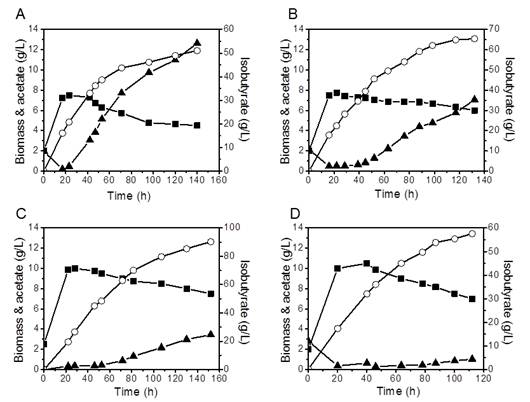
Figures(4)

Mingyong Xiong, Ping Yu, Jingyu Wang, Kechun Zhang. Improving Engineered Escherichia coli strains for High-level Biosynthesis of Isobutyrate[J]. AIMS Bioengineering, 2015, 2(2): 60-74. doi: 10.3934/bioeng.2015.2.60







 DownLoad:
DownLoad: