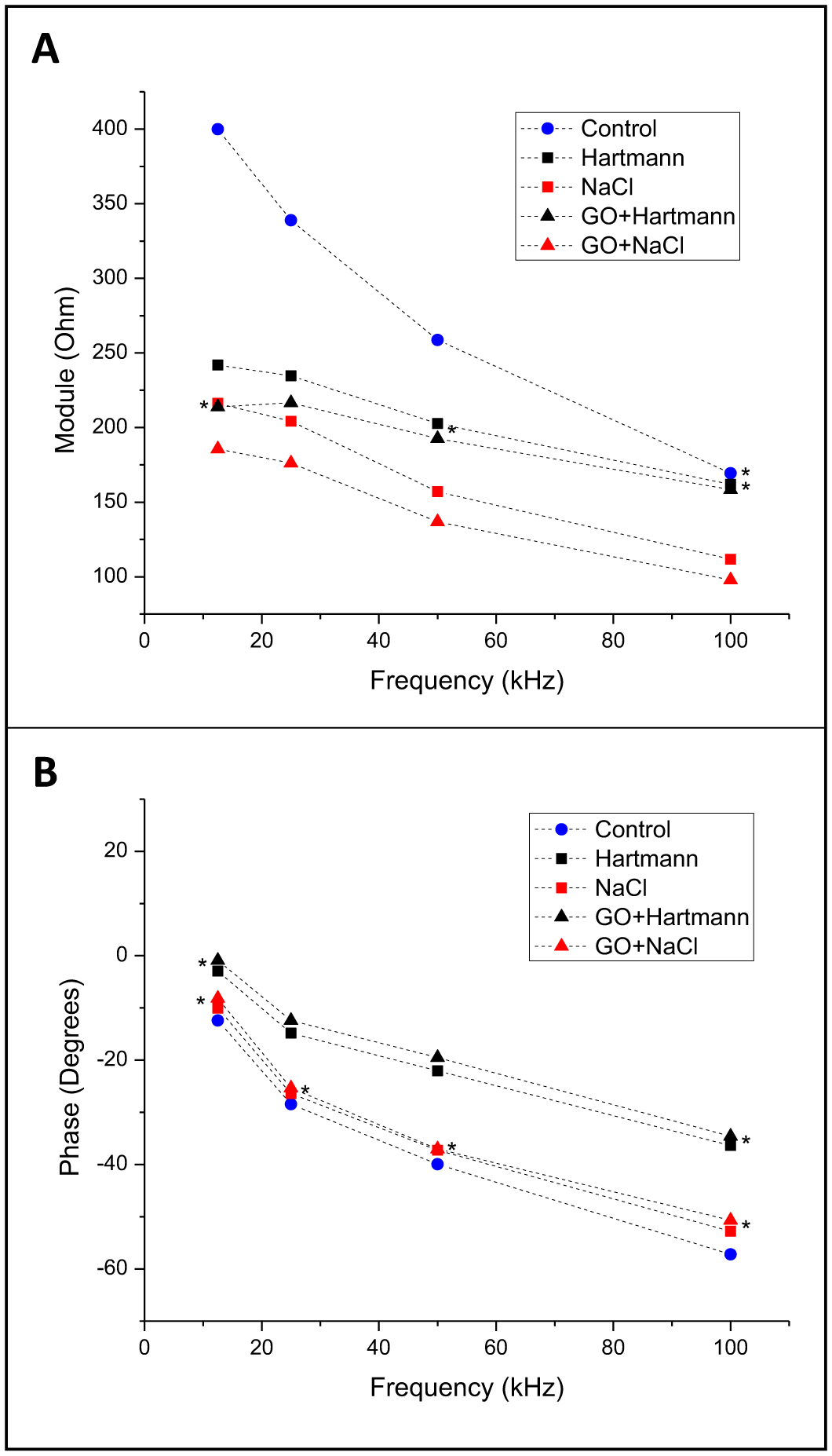
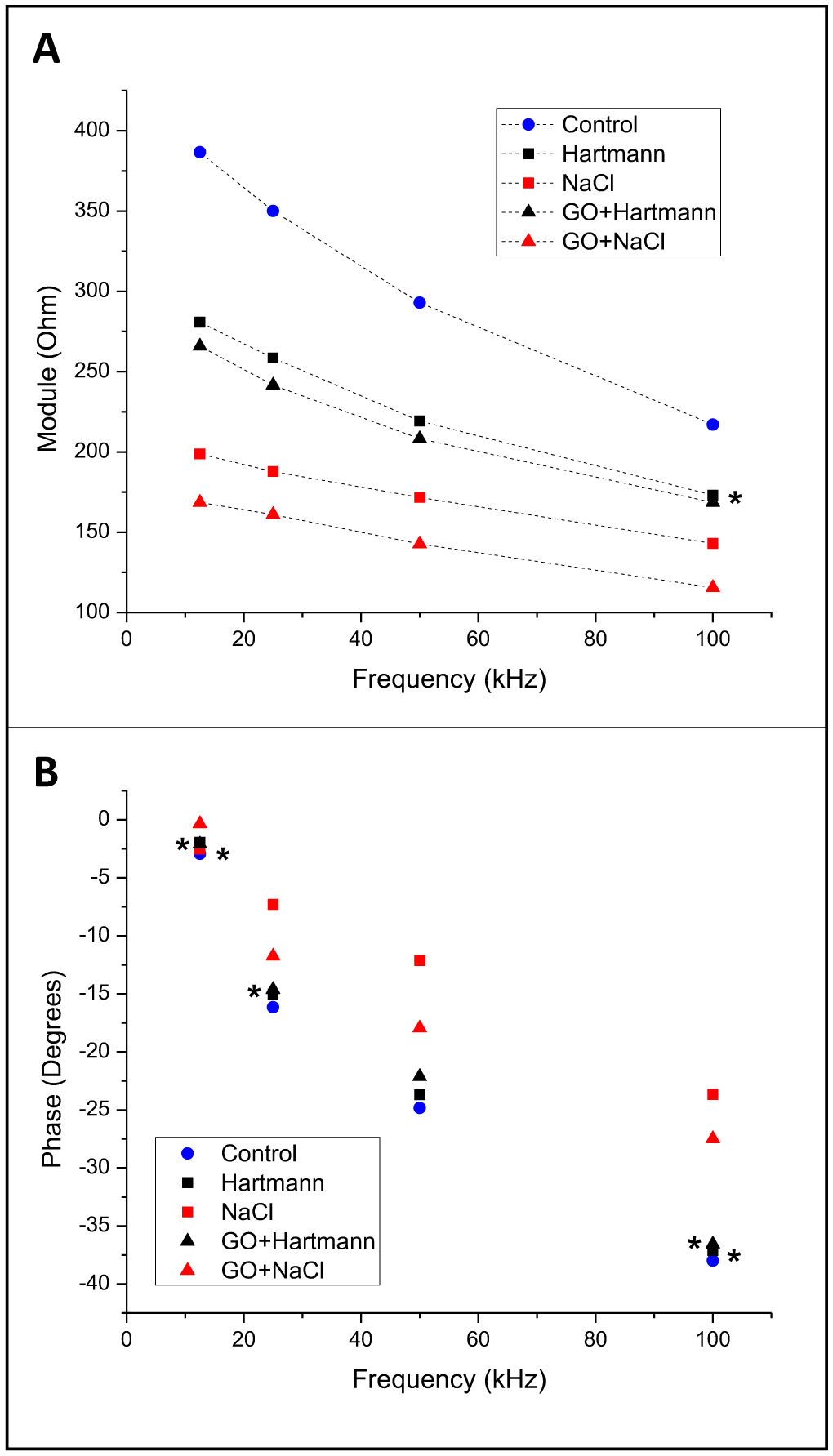
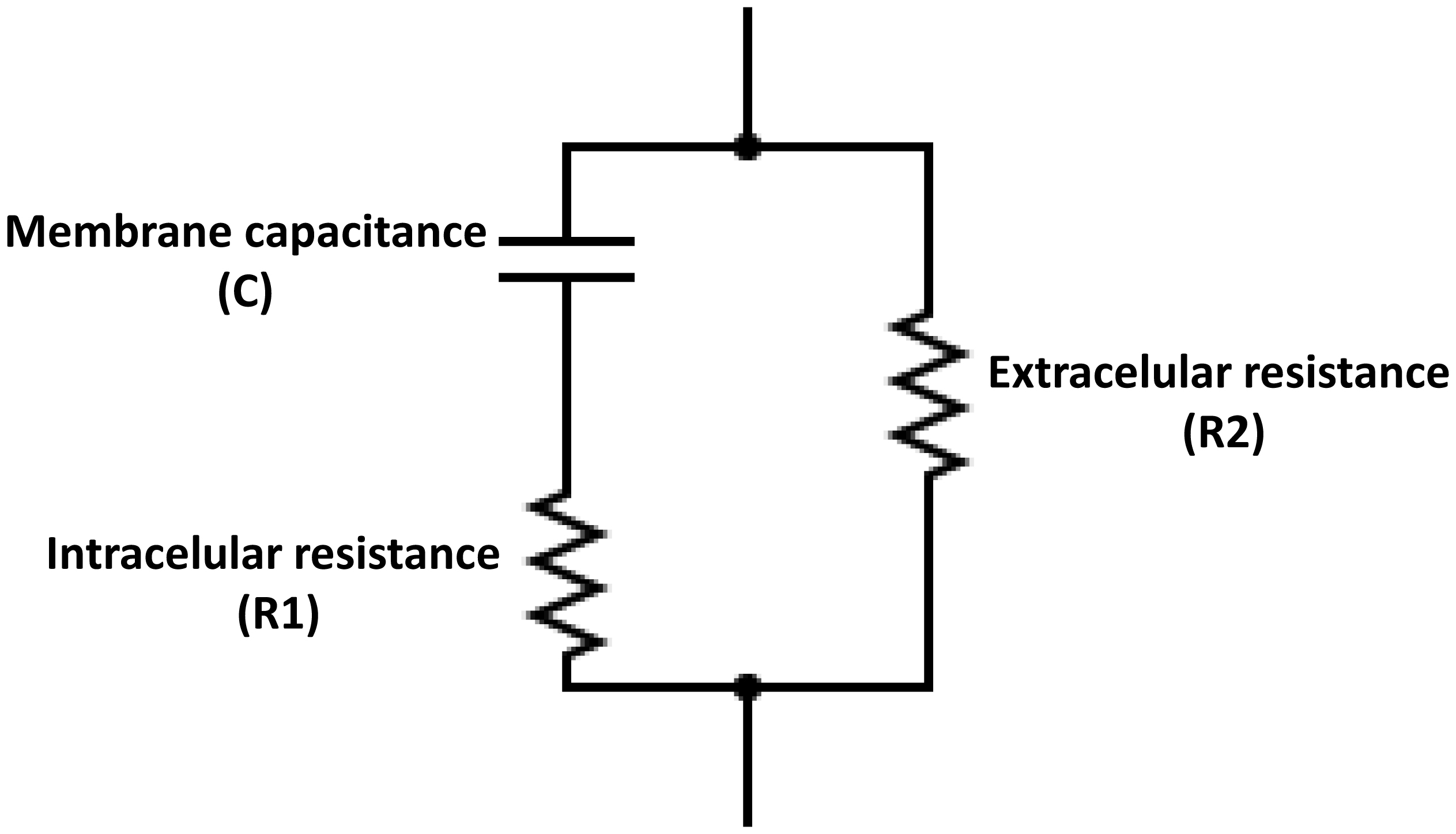
Electrical bioimpedance (BI) was proposed as an easy and cheap technique to monitor different physiological parameters. However, its clinical applications are minimal due to the low resolution and difficulties in discriminating between different tissue types. Nanoparticles have also been extensively investigated for their practical use. Graphene oxide (GO) has shown acceptable biocompatibility. The objective of this work was to assess the possibility of using GO to enhance bioimpedance measures in ex vivo models. GO was suspended in medical-grade solutions and injected into the tissues. BI was recorded at 12.5, 25, 50, and 100 kHz. Bode impedance plots evidenced statistically significant differences in tissue impedance before and after injections. Likewise, data adjustment to an equivalent electrical circuit showed that GO accumulates mainly in the extracellular space and, to some extent, at the cytoplasmatic membrane, and the accumulation is tissue specific. The latter suggests the possibility of using GO as a contrast agent to discriminate between different tissue types using BI.
Citation: Svetlana Kashina, Andrea Monserrat del Rayo Cervantes-Guerrero, Francisco Miguel Vargas-Luna, Gonzalo Paez, Jose Marco Balleza-Ordaz. Tissue-specific bioimpedance changes induced by graphene oxide ex vivo: a step toward contrast media development[J]. AIMS Biophysics, 2025, 12(1): 54-68. doi: 10.3934/biophy.2025005
Electrical bioimpedance (BI) was proposed as an easy and cheap technique to monitor different physiological parameters. However, its clinical applications are minimal due to the low resolution and difficulties in discriminating between different tissue types. Nanoparticles have also been extensively investigated for their practical use. Graphene oxide (GO) has shown acceptable biocompatibility. The objective of this work was to assess the possibility of using GO to enhance bioimpedance measures in ex vivo models. GO was suspended in medical-grade solutions and injected into the tissues. BI was recorded at 12.5, 25, 50, and 100 kHz. Bode impedance plots evidenced statistically significant differences in tissue impedance before and after injections. Likewise, data adjustment to an equivalent electrical circuit showed that GO accumulates mainly in the extracellular space and, to some extent, at the cytoplasmatic membrane, and the accumulation is tissue specific. The latter suggests the possibility of using GO as a contrast agent to discriminate between different tissue types using BI.

| [1] |
Simini F, Bertemes-Filho P (2018) Bioimpedance in Biomedical Applications and Research. New York: Springer. https://doi.org/10.1007/978-3-319-74388-2 
|
| [2] | Aldobali M, Pal K (2021) Bioelectrical impedance analysis for evaluation of body composition: a review. 2021 International Congress of Advanced Technology and Engineering (ICOTEN) : 1-10. https://doi.org/10.1109/ICOTEN52080.2021.9493494 |
| [3] |
Amini M, Hisdal J, Kalvøy H (2018) Applications of bioimpedance measurement techniques in tissue engineering. Electr Bioimpedance 9: 142-158. https://doi.org/10.2478/joeb-2018-0019
|
| [4] |
Lukaski HC, Vega Diaz N, Talluri A, et al. (2019) Classification of hydration in clinical conditions: indirect and direct approaches using bioimpedance. Nutrients 11: 809. https://doi.org/10.3390/nu11040809
|
| [5] |
Sanchez B, Vandersteen G, Martin I, et al. (2013) In vivo electrical bioimpedance characterization of human lung tissue during the bronchoscopy procedure. A feasibility study. Med Eng Phys 35: 949-957. https://doi.org/10.1016/j.medengphy.2012.09.004
|
| [6] |
Sanchez B, Louarroudi E, Jorge E, et al. (2013) A new measuring and identification approach for time-varying bioimpedance using multisine electrical impedance spectroscopy. Physiol Meas 34: 339. https://doi.org/10.1088/0967-3334/34/3/339
|
| [7] |
Subhan S, Ha S (2019) A harmonic error cancellation method for accurate clock-based electrochemical impedance spectroscopy. IEEE T Biomed Circ S 13: 710-724. https://doi.org/10.1109/TBCAS.2019.2923719
|
| [8] |
Schöckel L, Jost G, Seidensticker P, et al. (2020) Developments in X-ray contrast media and the potential impact on computed tomography. Invest Radiol 55: 592-597. https://doi.org/10.1097/RLI.0000000000000696
|
| [9] | Naranjo-Hernández D, Reina-Tosina J, Min M (2019) Fundamentals, recent advances, and future challenges in bioimpedance devices for healthcare applications. J Sensors 201: 9210258. https://doi.org/10.1155/2019/9210258 |
| [10] |
Sobota V, Müller M, Roubík K (2019) Intravenous administration of normal saline may be misinterpreted as a change of end-expiratory lung volume when using electrical impedance tomography. Sci Rep 9: 5775. https://doi.org/10.1038/s41598-019-42241-7
|
| [11] |
Putensen C, Hentze B, Muenster S, et al. (2019) Electrical impedance tomography for cardio-pulmonary monitoring. J Clin Med 8: 1176. https://doi.org/10.3390/jcm8081176
|
| [12] |
Cervantes A, Paez G, Balleza-Ordaz JM, et al. (2024) Electrical bioimpedance analysis and comparison in biological tissues through crystalloid solutions implementation. Biosens Bioelectron 246: 115874. https://doi.org/10.1016/j.bios.2023.115874
|
| [13] |
Avasthi A, Caro C, Pozo‑Torres E, et al. (2020) Magnetic nanoparticles as MRI contrast agents. Surface-modified Nanobiomaterials for Electrochemical and Biomedicine Applications . Cham: Springer 49-91. https://doi.org/10.1007/978-3-030-55502-3_3
|
| [14] |
Thakur A, Thakur P, Khurana SP (2022) Synthesis and Applications of Nanoparticles. Singapore: Springer. https://doi.org/10.1007/978-981-16-6819-7
|
| [15] |
Liao C, Li Y, Tjong SC (2018) Graphene nanomaterials: synthesis, biocompatibility, and cytotoxicity. Int J Mol Sci 19: 3564. https://doi.org/10.3390/ijms19113564
|
| [16] |
Sekhon SS, Kaur P, Kim YH, et al. (2021) 2D graphene oxide–aptamer conjugate materials for cancer diagnosis. Npj 2D Mater Appl 5: 21. https://doi.org/10.1038/s41699-021-00202-7
|
| [17] |
Huang A, Zhang L, Li W, et al. (2018) Controlled fluorescence quenching by antibody-conjugated graphene oxide to measure tau protein. Roy Soc Open Sci 5: 171808. https://doi.org/10.1098/rsos.171808
|
| [18] |
Campbell E, Hasan M, Pho C, et al. (2019) Graphene oxide as a multifunctional platform for intracellular delivery, imaging, and cancer sensing. Sci Rep 9: 416. https://doi.org/10.1038/s41598-018-36617-4
|
| [19] |
Casero E, Parra-Alfambra A, Petit-Domínguez M, et al. (2012) Differentiation between graphene oxide and reduced graphene by electrochemical impedance spectroscopy (EIS). Electrochem Commun 20: 63-66. https://doi.org/10.1016/j.elecom.2012.04.002
|
| [20] |
Yoon Y, Jo J, Kim S, et al. (2017) Impedance spectroscopy analysis and equivalent circuit modeling of graphene oxide solutions. Nanomaterials 7: 446. https://doi.org/10.3390/nano7120446
|
| [21] |
Magar HS, Hassan RY, Mulchandani A (2021) Electrochemical impedance spectroscopy (EIS): principles, construction, and biosensing applications. Sensors 21: 6578. https://doi.org/10.3390/s21196578
|
| [22] |
Chen P, Wang G, Li J, et al. (2024) Anticorrosion mechanism of FACs-GO hybrids in ER coatings by EIS and MD simulation. Prog Org Coat 186: 107996. https://doi.org/10.1016/j.porgcoat.2023.107996
|
| [23] |
Backes C, Abdelkader AM, Alonso C, et al. (2020) Production and processing of graphene and related materials. 2D Materials 7: 022001. https://doi.org/10.1088/2053-1583/ab1e0a
|
| [24] |
Lunney JK, Van Goor A, Walker KE, et al. (2021) Importance of the pig as a human biomedical model. Sci Transl Med 13: eabd5758. https://doi.org/10.1126/scitranslmed.abd5758
|
| [25] |
Abasi S, Aggas JR, Garayar-Leyva GG, et al. (2022) Bioelectrical impedance spectroscopy for monitoring mammalian cells and tissues under different frequency domains: a review. ACS Meas Sci Au 2: 495-516. https://doi.org/10.1021/acsmeasuresciau.2c00033
|
| [26] | Bera TK (2018) Bioelectrical impedance and the frequency dependent current conduction through biological tissues: a short review. IOP Conference Series: Materials Science and Engineering . |
| [27] |
Ferrari AC, Basko DM (2013) Raman spectroscopy as a versatile tool for studying the properties of graphene. Nat Nanotechnol 8: 235-246. https://doi.org/10.1038/nnano.2013.46
|
| [28] |
Frerichs I, Hinz J, Herrmann P, et al. (2002) Regional lung perfusion as determined by electrical impedance tomography in comparison with electron beam CT imaging. IEEE T Med Imag 21: 646-652. https://doi.org/10.1109/TMI.2002.800585
|
| [29] | Hellige NC, Meyer B, Rodt T, et al. (2012) In-vitro evaluation of contrast media for assessment of regional perfusion distribution by Electrical Impedance Tomography (EIT). Biomed Eng Biomed Te . https://doi.org/10.1515/bmt-2012-4442 |
| [30] |
Qu G, Wang X, Liu Q, et al. (2013) The ex vivo and in vivo biological performances of graphene oxide and the impact of surfactant on graphene oxid's biocompatibility. J Environ Sci 25: 873-881. https://doi.org/10.1016/S1001-0742(12)60252-6
|
| [31] |
Faes T, Van Der Meij H, De Munck J, et al. (1999) The electric resistivity of human tissues (100 Hz-10 MHz): a meta-analysis of review studies. Physiol Meas 20: R1. https://doi.org/10.1088/0967-3334/20/4/201
|
| [32] |
Luo Y, Abiri P, Zhang S, et al. (2018) Non-invasive electrical impedance tomography for multi-scale detection of liver fat content. Theranostics 8: 1636-1647. https://doi.org/10.7150/thno.22233
|
| [33] |
Jasim DA, Murphy S, Newman L, et al. (2016) The effects of extensive glomerular filtration of thin graphene oxide sheets on kidney physiology. ACS Nano 10: 10753-10767. https://doi.org/10.1021/acsnano.6b03358
|
| [34] |
Wei XF, Grill WM (2009) Impedance characteristics of deep brain stimulation electrodes in vitro and in vivo. J Neural Eng 6: 046008. https://doi.org/10.1088/1741-2560/6/4/046008
|
| [35] |
Crone C, Olesen S (1982) Electrical resistance of brain microvascular endothelium. Brain Res 241: 49-55. https://doi.org/10.1016/0006-8993(82)91227-6
|
| [36] |
Butt AM, Jones HC, Abbott NJ (1990) Electrical resistance across the blood-brain barrier in anaesthetized rats: a developmental study. J Physiol 429: 47-62. https://doi.org/10.1113/jphysiol.1990.sp018243
|

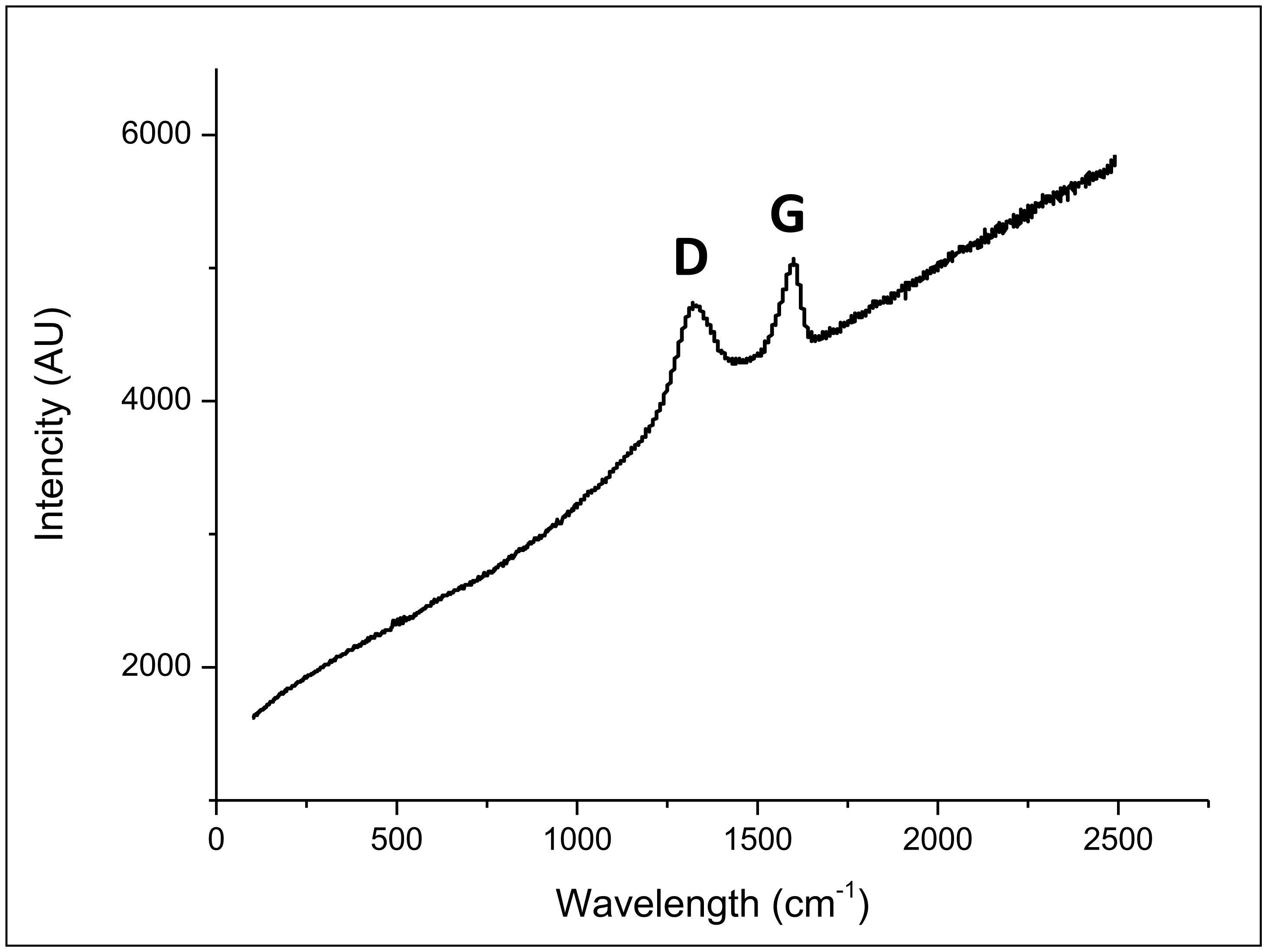
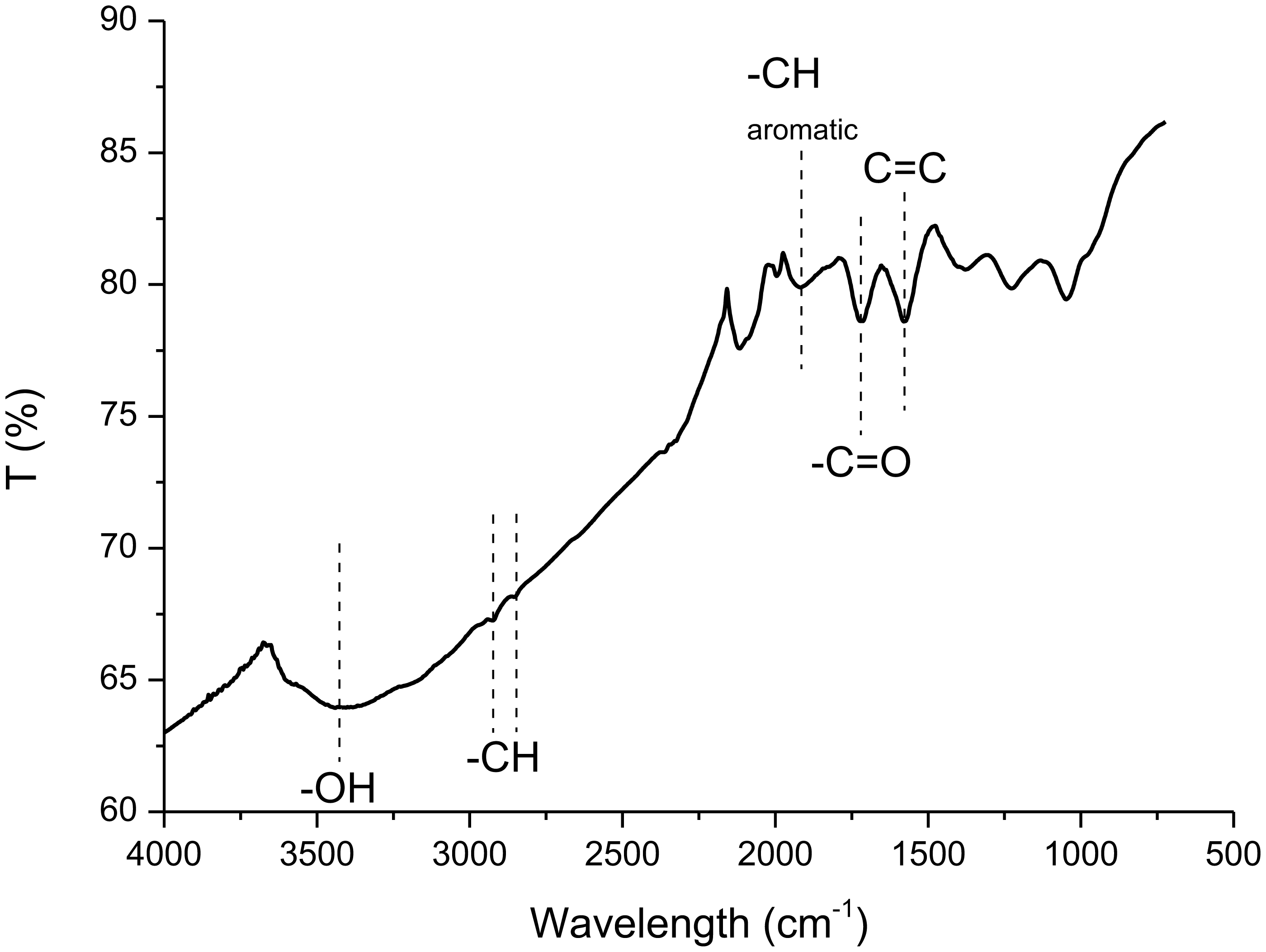
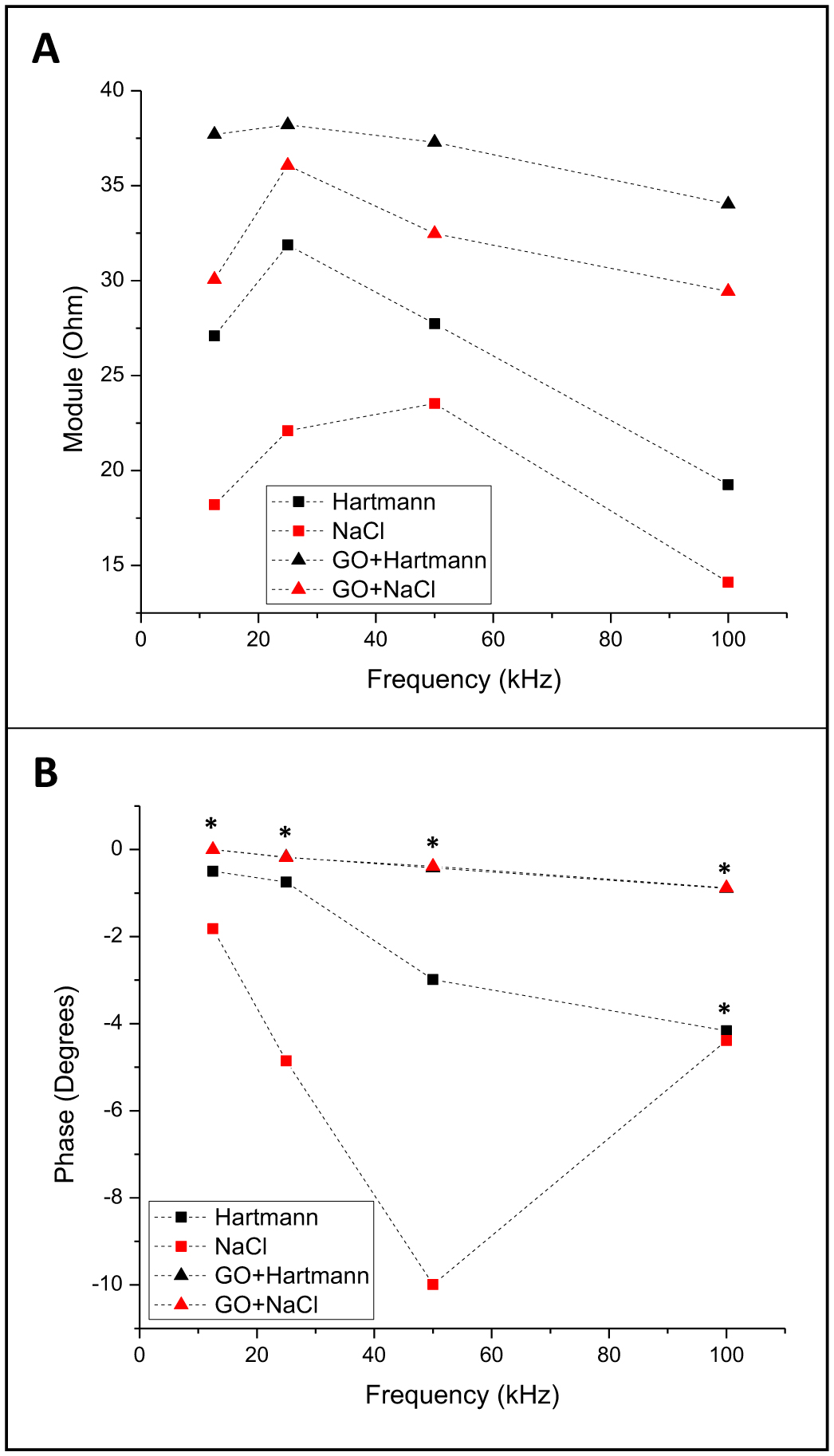
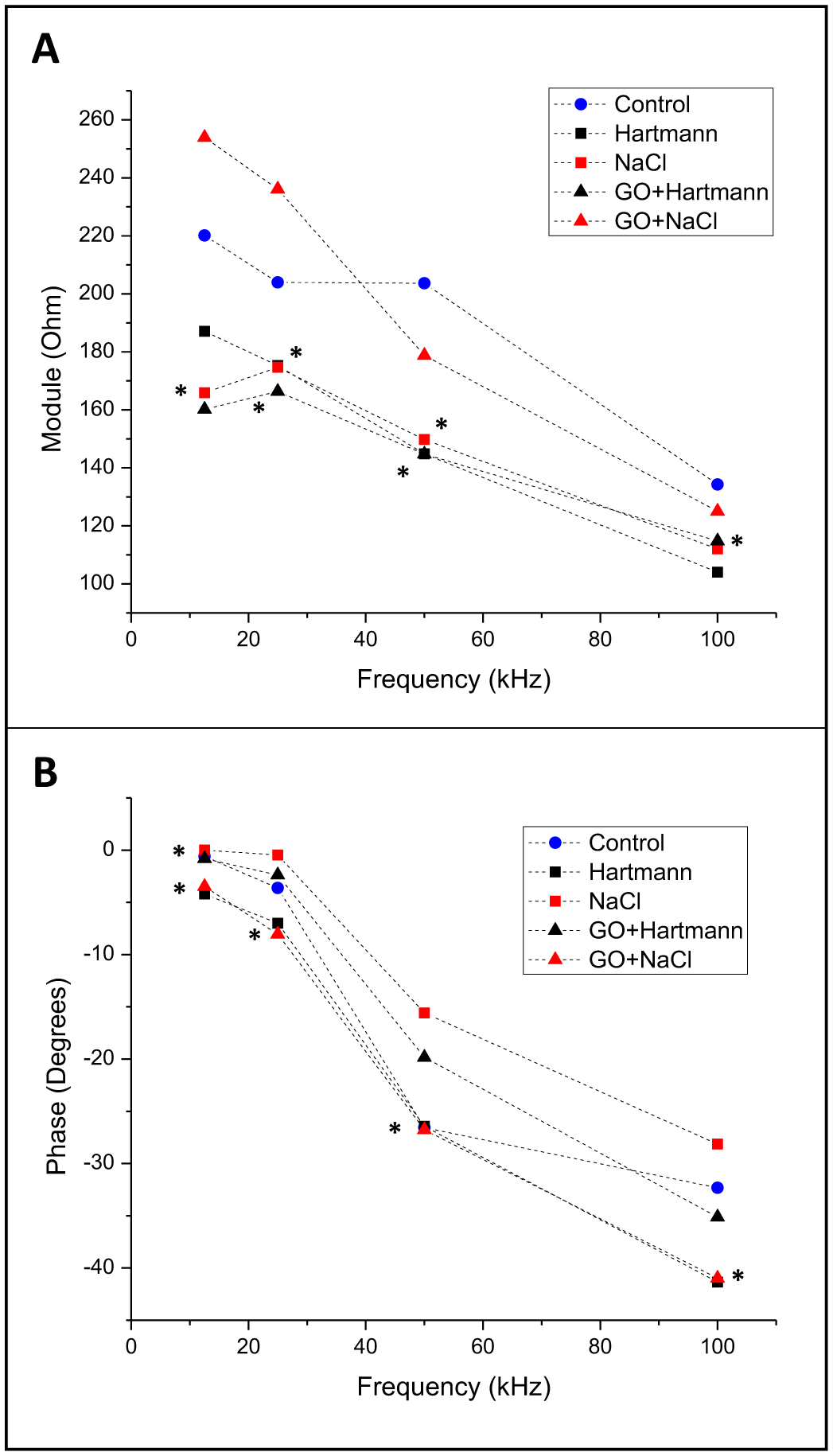
Figures(8) / Tables(1)

Svetlana Kashina, Andrea Monserrat del Rayo Cervantes-Guerrero, Francisco Miguel Vargas-Luna, Gonzalo Paez, Jose Marco Balleza-Ordaz. Tissue-specific bioimpedance changes induced by graphene oxide ex vivo: a step toward contrast media development[J]. AIMS Biophysics, 2025, 12(1): 54-68. doi: 10.3934/biophy.2025005







 DownLoad:
DownLoad: