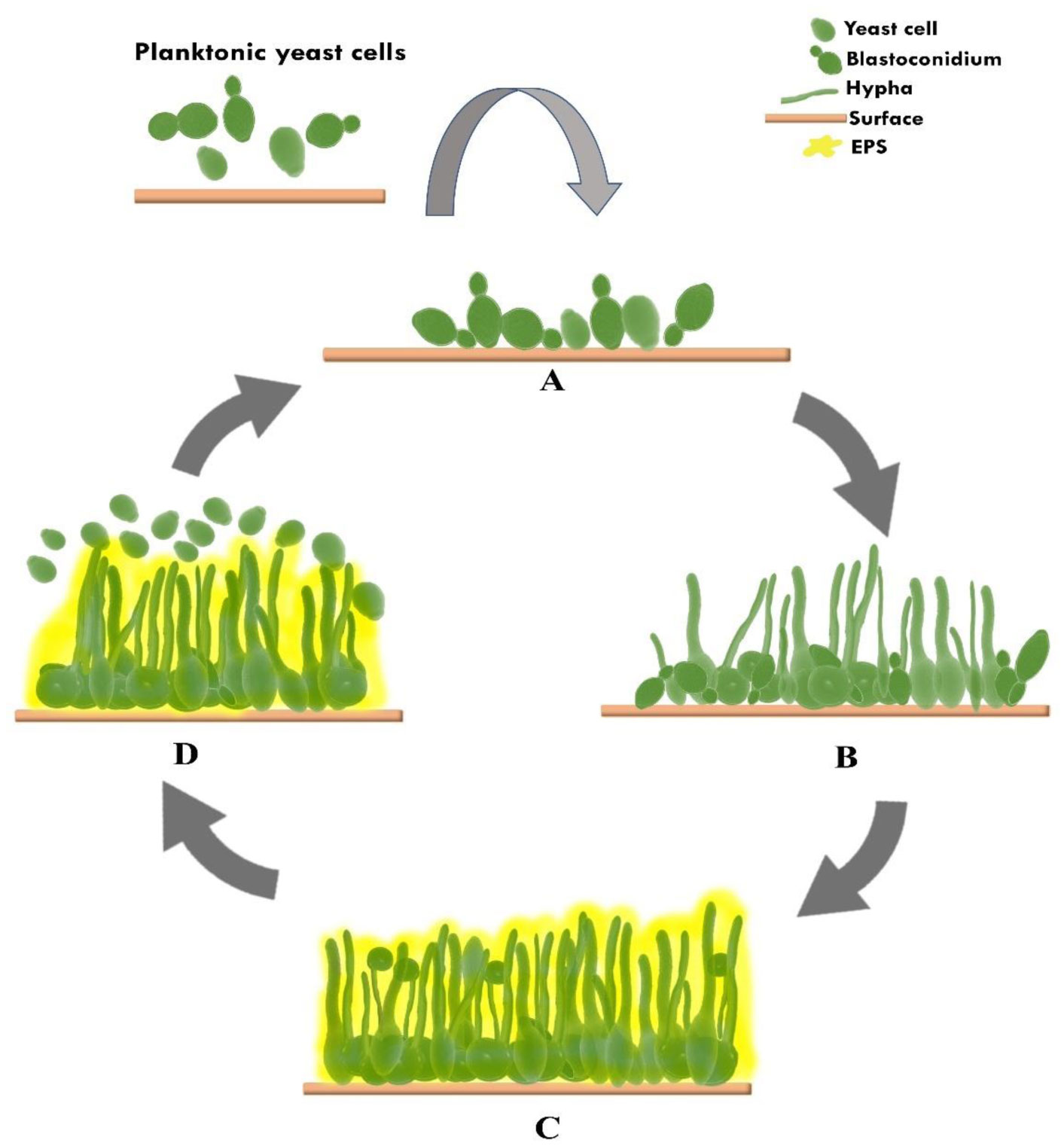
Candidemia is the most common form of invasive fungal infection associated with several risk factors, and one of them is the use of medical devices, to which microbial biofilms can attach. Candidemia related to the use of peripheral intravascular and central venous catheters (CVC) is referred to as Candida catheter-related bloodstream infection, with more than 90% being related to CVC usage. The infection is associated with a higher morbidity and mortality rate than nosocomial bacterial infections. Candida spp. can protect themselves from the host immune system and antifungal drugs because of the biofilm structure, which is potentiated by the extracellular matrix (ECM). Candida albicans and Candida parapsilosis are the most pathogenic species often found to form biofilms associated with catheter usage. Biofilm formation of C. albicans includes four mechanisms: attachment, morphogenesis, maturation and dispersion. The biofilms formed between C. albicans and non-albicans spp. differ in ECM structure and composition and are associated with the persistence of colonization to infection for various catheter materials and antifungal resistance. Efforts to combat Candida spp. biofilm formation on catheters are still challenging because not all patients, especially those who are critically ill, can be recommended for catheter removal; also to be considered are the characteristics of the biofilm itself, which readily colonizes the permanent medical devices used. The limited choice and increasing systemic antifungal resistance also make treating it more difficult. Hence, alternative strategies have been developed to manage Candida biofilm. Current options for prevention or therapy in combination with systemic antifungal medications include lock therapy, catheter coating, natural peptide products and photodynamic inactivation.
Citation: Meiliyana Wijaya, Ryan Halleyantoro, Jane Florida Kalumpiu. Biofilm: The invisible culprit in catheter-induced candidemia[J]. AIMS Microbiology, 2023, 9(3): 467-485. doi: 10.3934/microbiol.2023025
Candidemia is the most common form of invasive fungal infection associated with several risk factors, and one of them is the use of medical devices, to which microbial biofilms can attach. Candidemia related to the use of peripheral intravascular and central venous catheters (CVC) is referred to as Candida catheter-related bloodstream infection, with more than 90% being related to CVC usage. The infection is associated with a higher morbidity and mortality rate than nosocomial bacterial infections. Candida spp. can protect themselves from the host immune system and antifungal drugs because of the biofilm structure, which is potentiated by the extracellular matrix (ECM). Candida albicans and Candida parapsilosis are the most pathogenic species often found to form biofilms associated with catheter usage. Biofilm formation of C. albicans includes four mechanisms: attachment, morphogenesis, maturation and dispersion. The biofilms formed between C. albicans and non-albicans spp. differ in ECM structure and composition and are associated with the persistence of colonization to infection for various catheter materials and antifungal resistance. Efforts to combat Candida spp. biofilm formation on catheters are still challenging because not all patients, especially those who are critically ill, can be recommended for catheter removal; also to be considered are the characteristics of the biofilm itself, which readily colonizes the permanent medical devices used. The limited choice and increasing systemic antifungal resistance also make treating it more difficult. Hence, alternative strategies have been developed to manage Candida biofilm. Current options for prevention or therapy in combination with systemic antifungal medications include lock therapy, catheter coating, natural peptide products and photodynamic inactivation.

| [1] |
Cavalheiro M, Teixeira MC (2018) Candida biofilms: Threats, challenges, and promising strategies. Front Med 5: 28. https://doi.org/10.3389/fmed.2018.00028 
|
| [2] |
Pereira R, dos Santos Fontenelle RO, de Brito EHS, et al. (2021) Biofilm of Candida albicans: formation, regulation and resistance. J Appl Microbiol 131: 11-22. https://doi.org/10.1111/jam.14949
|
| [3] |
Atiencia-Carrera MB, Cabezas-Mera FS, Tejera E, et al. (2022) Prevalence of biofilms in Candida spp. bloodstream infections: A meta-analysis. PLoS One 17: e0263522. https://doi.org/10.1371/journal.pone.0263522
|
| [4] |
Pokhrel S, Boonmee N, Tulyaprawat O, et al. (2022) Assessment of biofilm formation by Candida albicans strains isolated from hemocultures and their role in pathogenesis in the zebrafish model. J Fungi 8: 1014. https://doi.org/10.3390/jof8101014
|
| [5] | Chen YN, Hsu JF, Chu SM, et al. (2022) Clinical and microbiological characteristics of neonates with candidemia and impacts of therapeutic strategies on the outcomes. J Fungi (Basel, Switzerland) 8: 465. https://doi.org/10.3390/jof8050465 |
| [6] |
Rodrigues LS, Motta FA, Picharski GL, et al. (2019) Invasive candidiasis: Risk factor for mortality in a pediatric tertiary care hospital in south of Brazil. Medicine (Baltimore) 98: e15933. https://doi.org/10.1097/MD.0000000000015933
|
| [7] |
Zhang W, Song X, Wu H, et al. (2019) Epidemiology, risk factors and outcomes of Candida albicans vs. non-albicans candidemia in adult patients in Northeast China. Epidemiol Infect 147: e277. https://doi.org/10.1017/S0950268819001638
|
| [8] |
Guinea J (2014) Global trends in the distribution of Candida species causing candidemia. Clin Microbiol Infect 20: 5-10. https://doi.org/10.1111/1469-0691.12539
|
| [9] |
Delaloye J, Calandra T (2014) Invasive candidiasis as a cause of sepsis in the critically ill patient. Virulence 5: 161-169. https://doi.org/10.4161/viru.26187
|
| [10] |
Bassetti M, Vena A, Meroi M, et al. (2020) Factors associated with the development of septic shock in patients with candidemia: A post hoc analysis from two prospective cohorts. Crit Care 24: 117. https://doi.org/10.1186/s13054-020-2793-y
|
| [11] |
Bassetti M, Vena A, Russo A (2018) Management of patients with septic shock due to Candida infection. Hosp Pract (1995) 46: 258-265. https://doi.org/10.1080/21548331.2018.1518104
|
| [12] | Kotey FC, Dayie NT, Tetteh-Uarcoo PB, et al. (2021) Candida bloodstream infections: changes in epidemiology and increase in drug resistance. Infect Dis Res Treat 14. https://doi.org/10.1177/11786337211026927 |
| [13] | Alves J, Alonso-Tarrés C, Rello J (2022) How to identify invasive candidemia in ICU-a narrative review. Antibiot (Basel, Switzerland) 11: 1804. https://doi.org/10.3390/antibiotics11121804 |
| [14] |
Bujdáková H (2016) Management of Candida biofilms: State of knowledge and new options for prevention and eradication. Future Microbiol 11: 235-251. https://doi.org/10.2217/fmb.15.139
|
| [15] |
Eix EF, Nett JE (2020) How biofilm growth affects Candida-host interactions. Front Microbiol 11: 1437. https://doi.org/10.3389/fmicb.2020.01437
|
| [16] |
Alenazy H, Alghamdi A, Pinto R, et al. (2021) Candida colonization as a predictor of invasive candidiasis in non-neutropenic ICU patients with sepsis: A systematic review and meta-analysis. Int J Infect Dis 102: 357-362. https://doi.org/10.1016/j.ijid.2020.10.092
|
| [17] |
Hu B, Du Z, Kang Y, et al. (2014) Catheter-related Candida bloodstream infection in intensive care unit patients: a subgroup analysis of the China-SCAN study. BMC Infect Dis 14: 594. https://doi.org/10.1186/s12879-014-0594-0
|
| [18] |
Pappas PG, Lionakis MS, Arendrup MC, et al. (2016) Invasive candidiasis. Nat Rev Dis Primers 4: 18026. https://doi.org/10.1038/nrdp.2018.26
|
| [19] |
Bouza E, Guinea J, Guembe M (2014) The role of antifungals against Candida biofilm in catheter-related candidemia. Antibiotics 4: 1-17. https://doi.org/10.3390/antibiotics4010001
|
| [20] |
Aldea Mansilla C, Martínez-Alarcón J, Gracia Ahufinger I, et al. (2019) Microbiological diagnosis of catheter-related infections. Enferm Infecc Microbiol Clin 37: 668-672. https://doi.org/10.1016/j.eimc.2018.07.009
|
| [21] |
Brunetti G, Navazio AS, Giuliani A, et al. (2019) Candida blood stream infections observed between 2011 and 2016 in a large Italian University Hospital: A time-based retrospective analysis on epidemiology, biofilm production, antifungal agents consumption and drug-susceptibility. PLoS One 14: e0224678. https://doi.org/10.1371/journal.pone.0224678
|
| [22] | Duggan S, Leonhardt I, Hünniger K, et al. (2015) Host response to Candida albicans bloodstream infection and sepsis. Virulence 6: 316-326. https://doi.org/10.4161/21505594.2014.988096 |
| [23] |
Panpetch W, Somboonna N, Bulan DE, et al. (2017) Oral administration of live- or heat-killed Candida albicans worsened cecal ligation and puncture sepsis in a murine model possibly due to an increased serum (1→3)-β-D-glucan. PLoS One 12: e0181439. https://doi.org/10.1371/journal.pone.0181439
|
| [24] |
Gulati M, Nobile CJ (2016) Candida albicans biofilms: development, regulation, and molecular mechanisms. Microbes Infect 18: 310-321. https://doi.org/10.1016/j.micinf.2016.01.002
|
| [25] |
Ponde NO, Lortal L, Ramage G, et al. (2021) Candida albicans biofilms and polymicrobial interactions. Crit Rev Microbiol 47: 91-111. https://doi.org/10.1080/1040841X.2020.1843400
|
| [26] |
Tsui C, Kong EF, Jabra-Rizk MA (2016) Pathogenesis of Candida albicans biofilm. Pathog Dis 74: ftw018. https://doi.org/10.1093/femspd/ftw018
|
| [27] |
D'Enfert C, Kaune AK, Alaban LR, et al. (2021) The impact of the fungus-host-microbiota interplay upon Candida albicans infections: current knowledge and new perspectives. FEMS Microbiol Rev 45: fuaa060. https://doi.org/10.1093/femsre/fuaa060
|
| [28] |
Uppuluri P, Acosta Zaldívar M, Anderson MZ, et al. (2018) Candida albicans dispersed cells are developmentally distinct from biofilm and planktonic cells. MBio 9. https://doi.org/10.1128/mBio.01338-18
|
| [29] |
Sachivkina N, Podoprigora I, Bokov D (2021) Morphological characteristics of Candida albicans, Candida krusei, Candida guilliermondii, and Candida glabrata biofilms, and response to farnesol. Vet World 14: 1608-1614. https://doi.org/10.14202/vetworld.2021.1608-1614
|
| [30] |
Desai JV, Mitchell AP (2015) Candida albicans biofilm development and its genetic control. Microbiol Spectr 3. https://doi.org/10.1128/microbiolspec.MB-0005-2014
|
| [31] |
Andes D, Nett J, Oschel P, et al. (2004) Development and characterization of an in vivo central venous catheter Candida albicans Biofilm Model. Infect Immun 72: 6023-6031. https://doi.org/10.1128/IAI.72.10.6023-6031.2004
|
| [32] |
Xu Z, Huang T, Min D, et al. (2022) Regulatory network controls microbial biofilm development, with Candida albicans as a representative: from adhesion to dispersal. Bioengineered 13: 253-267. https://doi.org/10.1080/21655979.2021.1996747
|
| [33] |
Araújo D, Henriques M, Silva S (2017) Portrait of Candida species biofilm regulatory network genes. Trends Microbiol 25: 62-75. https://doi.org/10.1016/j.tim.2016.09.004
|
| [34] |
McCall AD, Pathirana RU, Prabhakar A, et al. (2019) Candida albicans biofilm development is governed by cooperative attachment and adhesion maintenance proteins. NPJ Biofilms Microbiomes 5: 21. https://doi.org/10.1038/s41522-019-0094-5
|
| [35] |
Wuyts J, Van Dijck P, Holtappels M (2018) Fungal persister cells: the basis for recalcitrant infections?. PLoS Pathog 14: e1007301. https://doi.org/10.1371/journal.ppat.1007301
|
| [36] |
Priya A, Pandian SK (2020) Piperine impedes biofilm formation and hyphal morphogenesis of Candida albicans. Front Microbiol 11: 756. https://doi.org/10.3389/fmicb.2020.00756
|
| [37] |
Pierce CG, Vila T, Romo JA, et al. (2017) The Candida albicans biofilm matrix: composition, structure and function. J Fungi 3: 14. https://doi.org/10.3390/jof3010014
|
| [38] | Butassi E, Svetaz L, Carpinella MC, et al. (2021) Fungal biofilms as a valuable target for the discovery of natural products that cope with the resistance of medically important fungi-latest findings. Antibiot (Basel, Switzerland) 10: 1053. https://doi.org/10.3390/antibiotics10091053 |
| [39] |
Nett JE, Sanchez H, Cain MT, et al. (2010) Genetic basis of Candida biofilm resistance due to drug-sequestering matrix glucan. J Infect Dis 202: 171-175. https://doi.org/10.1086/651200
|
| [40] |
Taff HT, Mitchell KF, Edward JA, et al. (2013) Mechanisms of Candida biofilm drug resistance. Future Microbiol 8: 1325-1337. https://doi.org/10.2217/fmb.13.101
|
| [41] |
Vediyappan G, Rossignol T, D'Enfert C (2010) Interaction of Candida albicans biofilms with antifungals: transcriptional response and binding of antifungals to beta-glucans. Antimicrob Agents Chemother 54: 2096-2111. https://doi.org/10.1128/AAC.01638-09
|
| [42] |
Martins M, Uppuluri P, Thomas DP, et al. (2010) Presence of extracellular DNA in the Candida albicans biofilm matrix and its contribution to biofilms. Mycopathologia 169: 323-331. https://doi.org/10.1007/s11046-009-9264-y
|
| [43] |
Seneviratne CJ, Jin L, Samaranayake LP (2008) Biofilm lifestyle of Candida: A mini review. Oral Dis 14: 582-590. https://doi.org/10.1111/j.1601-0825.2007.01424.x
|
| [44] |
Zuza-Alves DL, Silva-Rocha WP, Chaves GM (2017) An update on Candida tropicalis based on basic and clinical approaches. Front Microbiol 8: 1927. https://doi.org/10.3389/fmicb.2017.01927
|
| [45] |
Bravo Ruiz G, Lorenz A (2021) What do we know about the biology of the emerging fungal pathogen of humans Candida auris?. Microbiol Res 242: 126621. https://doi.org/10.1016/j.micres.2020.126621
|
| [46] |
Negri M, Martins M, Henriques M, et al. (2010) Examination of potential virulence factors of Candida tropicalis clinical isolates from hospitalized patients. Mycopathologia 169: 175-182. https://doi.org/10.1007/s11046-009-9246-0
|
| [47] |
Estivill D, Arias A, Torres-Lana A, et al. (2011) Biofilm formation by five species of Candida on three clinical materials. J Microbiol Methods 86: 238-242. https://doi.org/10.1016/j.mimet.2011.05.019
|
| [48] |
West PT, Peters SL, Olm MR, et al. (2021) Genetic and behavioral adaptation of Candida parapsilosis to the microbiome of hospitalized infants revealed by in situ genomics, transcriptomics, and proteomics. Microbiome 9: 142. https://doi.org/10.1186/s40168-021-01085-y
|
| [49] | Miyake A, Gotoh K, Iwahashi J, et al. (2022) Characteristics of biofilms formed by C. parapsilosis causing an outbreak in a neonatal intensive care unit. J Fungi (Basel, Switzerland) 8: 700. https://doi.org/10.3390/jof8070700 |
| [50] |
Kim SK, El Bissati K, Mamoun CB (2006) Amino acids mediate colony and cell differentiation in the fungal pathogen Candida parapsilosis. Microbiology 152: 2885-2894. https://doi.org/10.1099/mic.0.29180-0
|
| [51] |
De Rose DU, Santisi A, Ronchetti MP, et al. (2021) Invasive Candida infections in neonates after major surgery: current evidence and new directions. Pathog 10: 319. https://doi.org/10.3390/pathogens10030319
|
| [52] |
Yamin DH, Husin A, Harun A (2021) Risk Factors of Candida parapsilosis catheter-related bloodstream infection. Front Public Heal 9: 631865. https://doi.org/10.3389/fpubh.2021.631865
|
| [53] | lotfali E (2019) Biofilm: structure, antifungal resistance, proteomics and genomics of Candida species. Open Access J Mycol Mycol Sci 2: 1-12. https://doi.org/10.23880/oajmms-16000108 |
| [54] |
Pristov KE, Ghannoum MA (2019) Resistance of Candida to azoles and echinocandins worldwide. Clin Microbiol Infect 25: 792-798. https://doi.org/10.1016/j.cmi.2019.03.028
|
| [55] |
Sullivan D, Coleman D (1998) Candida dubliniensis: Characteristics and identification. J Clin Microbiol 36: 329-334. https://doi.org/10.1128/JCM.36.2.329-334.1998
|
| [56] |
Sampath A, Weerasekera M, Dilhari A, et al. (2017) Comparison of duplex PCR and phenotypic analysis in differentiating Candida dubliniensis from Candida albicans from oral samples. AMB Express 7: 141. https://doi.org/10.1186/s13568-017-0435-9
|
| [57] |
Weber K, Schulz B, Ruhnke M (2010) The quorum-sensing molecule E, E-farnesol-Its variable secretion and its impact on the growth and metabolism of Candida species. Yeast 27: 727-739. https://doi.org/10.1002/yea.1769. PMID: 20641010
|
| [58] |
Sardi JCO, Scorzoni L, Bernardi T, et al. (2013) Candida species: Current epidemiology, pathogenicity, biofilm formation, natural antifungal products and new therapeutic options. J Med Microbiol 62: 10-24. https://doi.org/10.1099/jmm.0.045054-0
|
| [59] |
Henriques M, Azeredo J, Oliveira R (2006) Candida albicans and Candida dubliniensis: Comparison of biofilm formation in terms of biomass and activity. Br J Biomed Sci 63: 5-11. https://doi.org/10.1080/09674845.2006.11732712
|
| [60] |
Ramage G, Walle KV, Wickes BL, et al. (2001) Biofilm formation by Candida dubliniensis. J Clin Microbiol 39: 3234-3240. https://doi.org/10.1128/JCM.39.9.3234-3240.2001
|
| [61] |
Spivak ES, Hanson KE (2018) Candida auris: An emerging fungal pathogen. J Clin Microbiol 56. https://doi.org/10.1128/JCM.01588-17
|
| [62] |
Horton MV, Nett JE (2020) Candida auris infection and biofilm formation: going beyond the surface. Curr Clin Microbiol Reports 7: 51-56. https://doi.org/10.1007/s40588-020-00143-7
|
| [63] |
Chowdhary A, Sharma C, Meis JF (2017) Candida auris: a rapidly emerging cause of hospital-acquired multidrug-resistant fungal infections globally. PLoS Pathog 13: e1006290. https://doi.org/10.1371/journal.ppat.1006290
|
| [64] |
Sherry L, Ramage G, Kean R, et al. (2017) Biofilm-forming capability of highly virulent, multidrug-resistant Candida auris. Emerg Infect Dis 23: 328-331. https://doi.org/10.3201/eid2302.161320
|
| [65] |
Bapat PS, Nobile CJ (2021) Photodynamic therapy is effective against Candida auris biofilms. Front Cell Infect Microbiol 11: 713092. https://doi.org/10.3389/fcimb.2021.713092
|
| [66] |
Sánchez-Vargas LO, Estrada-Barraza D, Pozos-Guillen AJ, et al. (2013) Biofilm formation by oral clinical isolates of Candida species. Arch Oral Biol 58: 1318-1326. https://doi.org/10.1016/j.archoralbio
|
| [67] | Zuo XS, Liu Y, Cai X, et al. (2021) Association of different Candida species with catheter-related candidemia, and the potential antifungal treatments against their adhesion properties and biofilm-forming capabilities. J Clin Lab Anal 35: e23738. https://doi.org/10.1002/jcla.23738 |
| [68] |
Rodrigues CF, Rodrigues ME, Silva S, et al. (2017) Candida glabrata biofilms: How far have we come?. J Fungi 3: 11. https://doi.org/10.3390/jof3010011
|
| [69] |
D'Enfert C, Janbon G (2015) Biofilm formation in Candida glabrata: What have we learnt from functional genomics approaches?. FEMS Yeast Res 16: fov111. https://doi.org/10.1093/femsyr/fov111
|
| [70] |
Cavalheiro M, Pereira D, Formosa-Dague C, et al. (2021) From the first touch to biofilm establishment by the human pathogen Candida glabrata: a genome-wide to nanoscale view. Commun Biol 4: 886. https://doi.org/10.1038/s42003-021-02412-7
|
| [71] |
Gómez-Gaviria M, Mora-Montes HM (2020) Current aspects in the biology, pathogeny, and treatment of Candida krusei, a neglected fungal pathogen. Infect Drug Resist 13: 1673-1689. https://doi.org/10.2147/IDR.S247944
|
| [72] |
Pohl CH (2022) Recent advances and opportunities in the study of Candida albicans polymicrobial biofilms. Front Cell Infect Microbiol 12: 836379. https://doi.org/10.3389/fcimb.2022.836379
|
| [73] |
Rodrigues ME, Gomes F, Rodrigues CF (2019) Candida spp./bacteria mixed biofilms. J Fungi (Basel) 6: 5. https://doi.org/10.3390/jof6010005
|
| [74] |
Shirtliff ME, Peters BM, Jabra-Rizk MA (2009) Cross-kingdom interactions: Candida albicans and bacteria. FEMS Microbiol Lett 299: 1-8. https://doi.org/10.1111/j.1574-6968.2009.01668.x
|
| [75] |
Allison DL, Willems HME, Jayatilake JAMS, et al. (2016) Candida-bacteria interactions: their impact on human disease. Microbiol Spectr 4. https://doi.org/10.1128/microbiolspec.VMBF-0030-2016
|
| [76] |
Abrantes PM, Behardien K, Africa CW (2022) Real-time assessment of interspecies Candida biofilm formation. Open Microbiol J 16. https://doi.org/10.2174/18742858-v16-e221128-2022-26
|
| [77] |
Santos JD, Piva E, Vilela SF, et al. (2016) Mixed biofilms formed by C. albicans and non-albicans species: a study of microbial interactions. Braz Oral Res 30. https://doi.org/10.1590/1807-3107BOR-2016.vol30.0023
|
| [78] |
Rossoni RD, Barbosa JO, Vilela SF, et al. (2015) Competitive interactions between C. albicans, C. glabrata and C. krusei during biofilm formation and development of experimental candidiasis. PLoS One 10. https://doi.org/10.1371/journal.pone.0131700
|
| [79] |
Hosida TY, Cavazana TP, Henriques M, et al. (2018) Interactions between Candida albicans and Candida glabrata in biofilms: influence of the strain type, culture medium and glucose supplementation. Mycoses 61: 270-278. https://doi.org/10.1111/myc.12738
|
| [80] |
Pathirana RU, McCall AD, Norris HL, et al. (2019) Filamentous non-albicans Candida species adhere to Candida albicans and benefit from dual biofilm growth. Front Microbiol 10: 1188. https://doi.org/10.3389/fmicb.2019.01188
|
| [81] |
Barros PP, Ribeiro FC, Rossoni RD, et al. (2016) Influence of Candida krusei and Candida glabrata on Candida albicans gene expression in in vitro biofilms. Arch Oral Biol 64: 92-101. https://doi.org/10.1016/j.archoralbio.2016.01.005
|
| [82] |
Pappas PG, Kauffman CA, Andes DR, et al. (2015) Clinical practice guideline for the management of candidiasis: 2016 update by the infectious diseases society of America. Clin Infect Dis 62: e1-e50. https://doi.org/10.1093/cid/civ933
|
| [83] |
Imbert C, Rammaert B (2018) What could be the role of antifungal lock-solutions? From bench to bedside. Pathogens 7: 6. https://doi.org/10.3390/pathogens7010006
|
| [84] |
Martin-Loeches I, Antonelli M, Cuenca-Estrella M, et al. (2019) ESICM/ESCMID task force on practical management of invasive candidiasis in critically ill patients. Intensive Care Med 45: 789-805. https://doi.org/10.1007/s00134-019-05599-w
|
| [85] |
Basas J, Palau M, Gomis X, et al. (2019) Efficacy of liposomal amphotericin B and anidulafungin using an antifungal lock technique (ALT) for catheter-related Candida albicans and Candida glabrata infections in an experimental model. PLoS One 14: e0212426. https://doi.org/10.1371/journal.pone.0212426
|
| [86] |
Revie NM, Iyer KR, Robbins N, et al. (2018) Antifungal drug resistance: evolution, mechanisms and impact. Curr Opin Microbiol 45: 70-76. https://doi.org/10.1016/j.mib.2018.02.005
|
| [87] |
Berkow EL, Lockhart SR (2017) Fluconazole resistance in Candida species: a current perspective. Infect Drug Resist 10: 237-245. https://doi.org/10.2147/IDR.S118892
|
| [88] |
Cavassin FB, Baú-Carneiro JL, Vilas-Boas RR, et al. (2021) Sixty years of amphotericin b: an overview of the main antifungal agent used to treat invasive fungal infections. Infect Dis Ther 10: 115-147. https://doi.org/10.1007/s40121-020-00382-7
|
| [89] |
Kovács R, Majoros L (2022) Antifungal lock therapy: An eternal promise or an effective alternative therapeutic approach?. Lett Appl Microbiol 74: 851-862. https://doi.org/10.1111/lam.13653
|
| [90] |
Walraven CJ, Lee SA (2012) Antifungal lock therapy. Antimicrob Agents Chemother 57: 1-8. https://doi.org/10.1128/AAC.masthead.57-1
|
| [91] |
Fujimoto K, Takemoto K (2018) Efficacy of liposomal amphotericin B against four species of Candida biofilms in an experimental mouse model of intravascular catheter infection. J Infect Chemother 24: 958-964. https://doi.org/10.1016/j.jiac.2018.08.011
|
| [92] |
Simitsopoulou M, Kyrpitzi D, Velegraki A, et al. (2014) Caspofungin at catheter lock concentrations eradicates mature biofilms of Candida lusitaniae and Candida guilliermondii. Antimicrob Agents Chemother 58: 4953-4956. https://doi.org/10.1128/AAC.03117-14
|
| [93] |
Lown L, Peters BM, Walraven CJ, et al. (2016) An optimized lock solution containing micafungin, ethanol and doxycycline inhibits Candida albicans and mixed C. albicans-Staphyloccoccus aureus biofilms. PLoS One 11: e0159225. https://doi.org/10.1371/journal.pone.0159225
|
| [94] |
Piersigilli F, Auriti C, Bersani I, et al. (2014) Antifungal lock therapy with combined 70% ethanol and micafungin in a critically ill infant. Pediatr Infect Dis J 33: 419-420. https://doi.org/10.1097/INF.0000000000000116
|
| [95] |
Vera-González N, Shukla A (2020) Advances in biomaterials for the prevention and disruption of Candida biofilms. Front Microbiol 11: 538602. https://doi.org/10.3389/fmicb.2020.538602
|
| [96] |
Michl TD, Giles C, Cross AT, et al. (2017) Facile single-step bioconjugation of the antifungal agent caspofungin onto material surfaces: Via an epoxide plasma polymer interlayer. RSC Adv 7: 27678-27681. https://doi.org/10.1039/C7RA03897F
|
| [97] |
Coad BR, Lamont-Friedrich SJ, Gwynne L, et al. (2015) Surface coatings with covalently attached caspofungin are effective in eliminating fungal pathogens. J Mater Chem B 3: 8469-8476. https://doi.org/10.1039/c5tb00961h
|
| [98] |
Bachmann SP, VandeWalle K, Ramage G, et al. (2002) In vitro activity of caspofungin against Candida albicans biofilms. Antimicrob Agents Chemother 46: 3591-3596. https://doi.org/10.1128/AAC.46.11.3591-3596.2002
|
| [99] |
Griesser SS, Jasieniak M, Coad BR, et al. (2015) Antifungal coatings by caspofungin immobilization onto biomaterials surfaces via a plasma polymer interlayer. Biointerphases 10: 04A307. https://doi.org/10.1116/1.4933108
|
| [100] |
Naderi J, Giles C, Saboohi S, et al. (2019) Surface coatings with covalently attached anidulafungin and micafungin prevent Candida albicans biofilm formation. J Antimicrob Chemother 74: 360-364. https://doi.org/10.1093/jac/dky437
|
| [101] |
Vargas-Blanco D, Lynn A, Rosch J, et al. (2017) A pre-therapeutic coating for medical devices that prevents the attachment of Candida albicans. Ann Clin Microbiol Antimicrob 16: 41. https://doi.org/10.1186/s12941-017-0215-z
|
| [102] |
Oshiro KGN, Rodrigues G, Monges BED, et al. (2019) Bioactive peptides against fungal biofilms. Front Microbiol 10: 2169. https://doi.org/10.3389/fmicb.2019.02169
|
| [103] |
Raman N, Lee MR, Palecek SP, et al. (2014) Polymer multilayers loaded with antifungal β-peptides kill planktonic Candida albicans and reduce formation of fungal biofilms on the surfaces of flexible catheter tubes. J Control Release 191: 54-62. https://doi.org/10.1016/j.jconrel.2014.05.026
|
| [104] |
Garcia LGS, Guedes GM de M, da Silva MLQ, et al. (2018) Effect of the molecular weight of chitosan on its antifungal activity against Candida spp. in planktonic cells and biofilm. Carbohydr Polym 195: 662-669. https://doi.org/10.1016/j.carbpol.2018.04.091
|
| [105] |
Huq MA, Ashrafudoulla M, Parvez MAK, et al. (2022) Chitosan-coated polymeric silver and gold nanoparticles: biosynthesis, characterization and potential antibacterial applications: a review. Polymers (Basel) 14: 5302. https://doi.org/10.3390/polym14235302
|
| [106] |
Li B, Pan L, Zhang H, et al. (2021) Recent developments on using nanomaterials to combat Candida albicans. Front Chem 9: 813973. https://doi.org/10.3389/fchem.2021.813973
|
| [107] |
Shariati A, Didehdar M, Razavi S, et al. (2022) Natural compounds: A hopeful promise as an antibiofilm agent against Candida species. Front Pharmacol 13: 917787. https://doi.org/10.3389/fphar.2022.917787
|
| [108] |
Singla RK, Dubey AK (2019) Molecules and metabolites from natural products as inhibitors of biofilm in Candida spp. pathogens. Curr Top Med Chem 19: 2567-2578. https://doi.org/10.2174/1568026619666191025154834
|
| [109] |
Nett JE (2014) Future directions for anti-biofilm therapeutics targeting Candida. Expert Rev Anti Infect Ther 12: 375-382. https://doi.org/10.1586/14787210.2014.885838
|
| [110] |
Huang MC, Shen M, Huang YJ, et al. (2018) Photodynamic inactivation potentiates the susceptibility of antifungal agents against the planktonic and biofilm cells of Candida albicans. Int J Mol Sci 19: 434. https://doi.org/10.3390/ijms19020434
|
| [111] |
Souza SO, Raposo BL, Sarmento-Neto JF, et al. (2022) Photoinactivation of yeast and biofilm communities of Candida albicans mediated by ZnTnHex-2-PyP4+ Porphyrin. J Fungi 8: 556. https://doi.org/10.3390/jof8060556
|
| [112] | Štefánek M, Černáková L, Dekkerová J, et al. (2022) Photodynamic inactivation effectively eradicates Candida auris biofilm despite its interference with the upregulation of CDR1 and MDR1 Efflux Genes. J fungi (Basel, Switzerland) 8: 1137. https://doi.org/10.3390/jof8111137 |
| [113] |
Voit T, Cieplik F, Regensburger J, et al. (2021) Spatial distribution of a porphyrin-based photosensitizer reveals mechanism of photodynamic inactivation of Candida albicans. Front Med 8: 641244. https://doi.org/10.3389/fmed.2021.641244
|
| [114] |
Donnelly RF, McCarron PA, Tunney MM (2008) Antifungal photodynamic therapy. Microbiol Res 163: 1-12. https://doi.org/10.1016/j.micres.2007.08.001
|
| [115] |
Kato IT, Prates RA, Sabino CP, et al. (2013) Antimicrobial photodynamic inactivation inhibits Candida albicans virulence factors and reduces in vivo pathogenicity. Antimicrob Agents Chemother 57: 445-451. https://doi.org/10.1128/AAC.01451-12
|
| [116] |
Quiroga ED, Mora SJ, Alvarez MG, et al. (2016) Photodynamic inactivation of Candida albicans by a tetracationic tentacle porphyrin and its analogue without intrinsic charges in presence of fluconazole. Photodiagnosis Photodyn Ther 13: 334-340. https://doi.org/10.1016/j.pdpdt.2015.10.005
|
Figures(1) / Tables(2)

Meiliyana Wijaya, Ryan Halleyantoro, Jane Florida Kalumpiu. Biofilm: The invisible culprit in catheter-induced candidemia[J]. AIMS Microbiology, 2023, 9(3): 467-485. doi: 10.3934/microbiol.2023025







 DownLoad:
DownLoad: