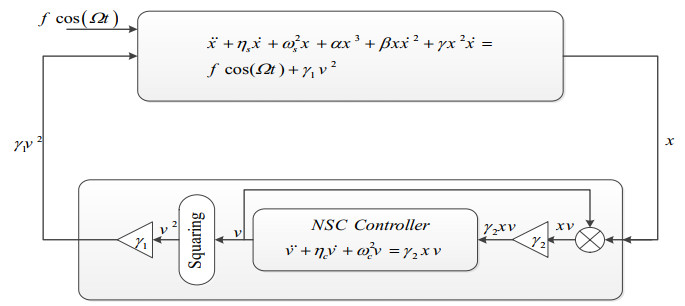
This paper studies the nonlinear vibrating behaviour of a nonlinear cantilever beam system (primary system) using a nonlinear absorber (the secondary system). The nonlinear vibrating behavior for the present dynamical system is considered with the effect of the external force. The one controller type, nonlinear saturation controller (NSC), is introduced to decrease the vibration of this system. Perturbation method treatment is produced to get the mathematical solution of the equations for the dynamical modeling with NSC. The perturbation technique is used to obtain the approximate solution of the dynamical system. This research focuses on resonance case with primary and 1:2 internal resonance. Time histories of the primary system and the controller are shown to demonstrate the reaction with and without control. The time-history response, as well as the impacts of the parameters on the system and controller, are simulated numerically using the MATLAB program. Routh-Hurwitz criterion is used to examine the stability of the system under primary resonance. A numerical simulation, using the MATLAB program, is obtained to show the time-history response, the effect of the parameters on the system and the controller. The effects of system parameters on the performance of the primary system and the controller are investigated. A comparison between all the obtained solutions made to confirm the results. Validation curves are provided to show how closely the perturbation and numerical solutions are related. A comparison is made with recently released papers.
Citation: Hany Bauomy, Ashraf Taha. Nonlinear saturation controller simulation for reducing the high vibrations of a dynamical system[J]. Mathematical Biosciences and Engineering, 2022, 19(4): 3487-3508. doi: 10.3934/mbe.2022161
This paper studies the nonlinear vibrating behaviour of a nonlinear cantilever beam system (primary system) using a nonlinear absorber (the secondary system). The nonlinear vibrating behavior for the present dynamical system is considered with the effect of the external force. The one controller type, nonlinear saturation controller (NSC), is introduced to decrease the vibration of this system. Perturbation method treatment is produced to get the mathematical solution of the equations for the dynamical modeling with NSC. The perturbation technique is used to obtain the approximate solution of the dynamical system. This research focuses on resonance case with primary and 1:2 internal resonance. Time histories of the primary system and the controller are shown to demonstrate the reaction with and without control. The time-history response, as well as the impacts of the parameters on the system and controller, are simulated numerically using the MATLAB program. Routh-Hurwitz criterion is used to examine the stability of the system under primary resonance. A numerical simulation, using the MATLAB program, is obtained to show the time-history response, the effect of the parameters on the system and the controller. The effects of system parameters on the performance of the primary system and the controller are investigated. A comparison between all the obtained solutions made to confirm the results. Validation curves are provided to show how closely the perturbation and numerical solutions are related. A comparison is made with recently released papers.

| [1] |
S. S. Oueini, A. H. Nayfeh, J. R. Pratt, A nonlinear vibration absorber for flexible structures, Nonlinear Dyn., 15 (1998), 259–282. https://doi.org/10.1023/A:1008250524547 doi: 10.1023/A:1008250524547

|
| [2] |
P. F. Pai, B. Wen, A. S. Naser, M. J. Schulz, Structural vibration control using PZT patches and non-linear phenomena, J. Sound Vib., 215 (1998), 273–296. https://doi.org/10.1006/jsvi.1998.1612 doi: 10.1006/jsvi.1998.1612
|
| [3] |
S. S. Oueini, A. H. Nayfeh, Single-mode control of a cantilever beam under principal parametric excitation, J. Sound Vib., 224 (1999), 33–47. https://doi.org/10.1006/jsvi.1998.2028 doi: 10.1006/jsvi.1998.2028
|
| [4] |
P. F. Pai, M. J. Schulz, A refined nonlinear vibration absorber, Int. J. Mech. Sci., 42 (2000), 537–560. https://doi.org/10.1016/S0020-7403(98)00135-0 doi: 10.1016/S0020-7403(98)00135-0
|
| [5] |
P. F. Pai, B. Rommel, M. J. Schulz, Non-linear vibration absorbers using higher order internal resonances, J. Sound Vib., 234 (2000), 799–817. https://doi.org/10.1006/jsvi.1999.2896 doi: 10.1006/jsvi.1999.2896
|
| [6] |
O. N. Ashour, A. H. Nayfeh, Adaptive control of flexible structures using a nonlinear vibration absorber, Nonlinear Dyn., 28 (2002), 309–322. https://doi.org/10.1023/A:1015622630382 doi: 10.1023/A:1015622630382
|
| [7] |
L. Jun, H. X. Hua, R. Y. Shen, Saturation-based active absorber for a non-linear plant to a principal external excitation, Mech. Syst. Signal Process., 21 (2007), 1489–1498. https://doi.org/10.1016/j.ymssp.2006.03.001 doi: 10.1016/j.ymssp.2006.03.001
|
| [8] |
L. Jun, X. B. Li, H. X. Hua, Active nonlinear saturation-based control for suppressing the free vibration of a self-excited plant, Commun. Nonlinear Sci. Numer. Simul., 15 (2010), 1071–1079. https://doi.org/10.1016/j.cnsns.2009.05.028 doi: 10.1016/j.cnsns.2009.05.028
|
| [9] |
J. Xu, K. W. Chung, Y. Y. Zhao, Delayed saturation controller for vibration suppression in a stainless-steel beam, Nonlinear Dyn., 62 (2010), 177–193. https://doi.org/10.1007/s11071-010-9708-4 doi: 10.1007/s11071-010-9708-4
|
| [10] |
A. T. EL-Sayed, Resonance behavior in coupled Van der Pol harmonic oscillators with controllers and delayed feedback, J. Vib. Control, 27 (2020), 1155–1170. https://doi.org/10.1177/1077546320938182 doi: 10.1177/1077546320938182
|
| [11] |
J. Warminski, M. Bochenski, W. Jarzyna, P. Filipek, M. Augustyniak, Active suppression of nonlinear composite beam vibrations by selected control algorithms, Commun. Nonlinear Sci. Numer. Simul., 16 (2011), 2237–2248. https://doi.org/10.1016/j.cnsns.2010.04.055 doi: 10.1016/j.cnsns.2010.04.055
|
| [12] |
N. A. Saeed, W. A. El-Ganini, M. Eissa, Nonlinear time delay saturation-based controller for suppression of nonlinear beam vibrations, Appl. Math. Modell., 37 (2013), 8846–8864. https://doi.org/10.1016/j.apm.2013.04.010 doi: 10.1016/j.apm.2013.04.010
|
| [13] | Y. S. Hamed, S. K. Elagan, On the vibration behavior study of a nonlinear flexible composite beam under excitation forces via nonlinear active vibration controller, Int. J. Basic Appl. Sci., 13 (2013), 9–18. |
| [14] |
Y. S. Hamed, Y. A. Amer, Nonlinear saturation controller for vibration supersession of a nonlinear composite beam, J. Mech. Sci. Technol., 28 (2014), 2987–3002. https://doi.org/10.1007/s12206-014-0706-1 doi: 10.1007/s12206-014-0706-1
|
| [15] |
M. Kamel, A. Kandil, W. A. El-Ganaini, M. Eissa, Active vibration control of a nonlinear magnetic levitation system via Nonlinear Saturation Controller (NSC), Nonlinear Dyn., 77 (2014), 605–619. https://doi.org/10.1007/s11071-014-1323-3 doi: 10.1007/s11071-014-1323-3
|
| [16] |
E. Omidi, S. N. Mahmoodi, Sensitivity analysis of the nonlinear integral positive position feedback and integral resonant controllers on vibration suppression of nonlinear oscillatory systems, Commun. Nonlinear Sci. Numer. Simul., 22 (2015), 149–166. https://doi.org/10.1016/j.cnsns.2014.10.011 doi: 10.1016/j.cnsns.2014.10.011
|
| [17] |
J. C. Ji, N. Zhang, Suppression of the primary resonance vibrations of a forced nonlinear system using a dynamic vibration absorber, J. Sound Vib., 329 (2010), 2044–2056. https://doi.org/10.1016/j.jsv.2009.12.020 doi: 10.1016/j.jsv.2009.12.020
|
| [18] |
J. C. Ji, N. Zhang, Suppression of super-harmonic resonance response using a linear vibration absorber, Mech. Res. Commun., 38 (2011), 411–416. https://doi.org/10.1016/j.mechrescom.2011.05.014 doi: 10.1016/j.mechrescom.2011.05.014
|
| [19] |
J. C. Ji, Application of a weakly nonlinear absorber to suppress the resonant vibrations of a forced nonlinear oscillator, J. Vib. Acoust., 134 (2012), 044502. https://doi.org/10.1115/1.4005839 doi: 10.1115/1.4005839
|
| [20] |
J. C. Ji, Design of a nonlinear vibration absorber using three-to-one internal resonances, Mech. Syst. Signal Process., 42 (2014), 236–246. https://doi.org/10.1016/j.ymssp.2013.06.019 doi: 10.1016/j.ymssp.2013.06.019
|
| [21] |
H. S. Bauomy, A. T. El-Sayed, A new six-degrees of freedom model designed for a composite plate through PPF controllers, Appl. Math. Modell., 88 (2020), 604–630. https://doi.org/10.1016/j.apm.2020.06.067 doi: 10.1016/j.apm.2020.06.067
|
| [22] |
H. S. Bauomy, A. T. El-Sayed, Act of nonlinear proportional derivative controller for MFC laminated shell, Phys. Scr., 95 (2020), 095210. https://doi.org/10.1088/1402-4896/abaa7c doi: 10.1088/1402-4896/abaa7c
|
| [23] |
H. S. Bauomy, A. T. El-Sayed, Mixed controller (IRC+NSC) involved in the harmonic vibration response cantilever beam model, Meas. Control, 53 (2020), 1954–1967. https://doi.org/10.1177/0020294020964243 doi: 10.1177/0020294020964243
|
| [24] |
A. T. El-Sayed, H. S. Bauomy, Outcome of special vibration controller techniques linked to a cracked beam, Appl. Math. Modell., 63 (2018), 266–287. https://doi.org/10.1016/j.apm.2018.06.045 doi: 10.1016/j.apm.2018.06.045
|
| [25] |
H. S. Bauomy, New controller (NPDCVF) outcome of FG cylindrical shell structure, Alexandria Eng. J., 61 (2021), 1779–1801. https://doi.org/10.1016/j.aej.2021.06.061 doi: 10.1016/j.aej.2021.06.061
|
| [26] |
J. C. Ji, X. Y. Li, L. Z. Zhang, Two–to–one resonant Hopf bifurcation in a quadratically nonlinear oscillator involving time delay, Int. J. Bifurcation Chaos, 22 (2012), 1250060. https://doi.org/10.1142/S0218127412500605 doi: 10.1142/S0218127412500605
|
| [27] |
J. C. Ji, Secondary resonances of a quadratic nonlinear oscillator following two-to-one resonant Hopf bifurcations, Nonlinear Dyn., 78 (2014), 2161–2184. https://doi.org/10.1007/s11071-014-1588-6 doi: 10.1007/s11071-014-1588-6
|
| [28] |
W. Zhang, J. Li, Global analysis for a nonlinear vibration absorber with fast and slow modes, Int. J. Bifurcation Chaos, 11 (2001), 2179–2194. https://doi.org/10.1142/S0218127401003334 doi: 10.1142/S0218127401003334
|
| [29] |
W. Zhang, Chaotic motion and its control for nonlinear nonplanar oscillations of a parametrically excited cantilever beam, Chaos, Solitons Fractals, 26 (2005), 731–745. https://doi.org/10.1016/j.chaos.2005.01.042 doi: 10.1016/j.chaos.2005.01.042
|
| [30] |
W. Zhang, Z. Yao, M. Yao, Periodic and chaotic dynamics of laminated composite piezoelectric rectangular plate with one-to-two internal resonance, Sci. China, Ser. E: Technol. Sci., 52 (2009), 731–742. https://doi.org/10.1007/s11431-009-0051-2 doi: 10.1007/s11431-009-0051-2
|
| [31] |
Y. X. Hao, W. Zhang, J. Yang, Nonlinear oscillations of a cantilever FGM rectangular plate based on third-order plate theory and asymptotic perturbation method, Composites, Part B, 42 (2011), 402–413. https://doi.org/10.1016/j.compositesb.2010.12.010 doi: 10.1016/j.compositesb.2010.12.010
|
| [32] |
W. Zhang, M. H. Zhao, X. Y. Guo, Nonlinear responses of a symmetric cross-ply composite laminated cantilever rectangular plate under in-plane and moment excitations, Compos. Struct., 100 (2013), 554–565. https://doi.org/10.1016/j.compstruct.2013.01.013 doi: 10.1016/j.compstruct.2013.01.013
|
| [33] |
X. Y. Guo, W. Zhang, Nonlinear vibrations of a reinforced composite plate with carbon nanotubes, Compos. Struct., 135 (2016), 96–108. https://doi.org/10.1016/j.compstruct.2015.08.063 doi: 10.1016/j.compstruct.2015.08.063
|
| [34] |
S. F. Lu, Y. Jiang, W. Zhang, X. J. Song, Vibration suppression of cantilevered piezoelectric laminated composite rectangular plate subjected to aerodynamic force in hygrothermal environment, Eur. J. Mech. - A/Solids, 83 (2020), 104002. https://doi.org/10.1016/j.euromechsol.2020.104002 doi: 10.1016/j.euromechsol.2020.104002
|
| [35] |
W. Zhang, G. Liu, B. Siriguleng, Saturation phenomena and nonlinear resonances of rotating pretwisted laminated composite blade under subsonic air flow excitation, J. Sound Vib., 478 (2020), 115353. https://doi.org/10.1016/j.jsv.2020.115353 doi: 10.1016/j.jsv.2020.115353
|
| [36] | A. H. Nayfeh, Perturbation Methods, Wiley, New York, 2000. https://doi.org/10.1002/9783527617609 |
| [37] | A. H. Nayfeh, D. Mook, Nonlinear Oscillations, Wiley, New York, 1995. https://doi.org/10.1002/9783527617586 |















Figures(16)

Hany Bauomy, Ashraf Taha. Nonlinear saturation controller simulation for reducing the high vibrations of a dynamical system[J]. Mathematical Biosciences and Engineering, 2022, 19(4): 3487-3508. doi: 10.3934/mbe.2022161







 DownLoad:
DownLoad: