Citation: Soraya L. Sá Canabarro, Ana Garcia, Corina Satler, Maria Clotilde Henriques Tavares. Interaction between Neural and Cardiac Systems during the Execution of the Stroop Task by Young Adults: Electroencephalographic Activity and Heart Rate Variability[J]. AIMS Neuroscience, 2017, 4(1): 28-51. doi: 10.3934/Neuroscience.2017.1.28

| [1] | Squire LR, Berg D, Bloom FE, et al. (2008) The Prefrontal Cortex and Executive Brain Functions. Fundament Neurosci. 3rd Edition ed. London: Academic Press. pp. 1199-1222. |
| [2] |
Diamond A (2013) Executive Functions. Annu Rev Psychol 64: 135-168. doi: 10.1146/annurev-psych-113011-143750

|
| [3] |
Ardila A (2008) On the Evolutionary Origins of Executive Functions. Brain Cognition 68: 92-99. doi: 10.1016/j.bandc.2008.03.003
|
| [4] |
Chan RCK, Shum D, Toulopoulou T, et al. (2008) Assessment of Executive Functions: Review of Instruments and Identification of Critical Issues. Arc Clinical Neuropsychol 23: 201-216. doi: 10.1016/j.acn.2007.08.010
|
| [5] |
Elliot R (2003) Executive Functions and Their Disorders. British Medical Bulletin 65: 49-59. doi: 10.1093/bmb/65.1.49
|
| [6] |
Sauseng P, Gerloff C, Hummel F (2013) Two Brakes Are Better Than One: The Neural Bases of Inhibitory Control of Motor Memory Traces. NeuroImage 65: 52-58. doi: 10.1016/j.neuroimage.2012.09.048
|
| [7] | Bunge SA, Mackey AP, Whitaker KJ (2009) Brain Changes Underlying the Development of Cognitive Control and Reasoning. In: Gazzaniga MS, editor. The cognitive neurosciences. 4ª ed. ed. Cambridge, Massachusetts: The MIT Press. pp. 73-85. |
| [8] | Stroop JR (1935) Studies of Interference in Serial Verbal Reaction. J Experiment Psychol XVIII: 643-662. |
| [9] |
Van der Elst W, Van Boxtel MPJ, Van Breukelen GJP, et al. (2006) The Stroop Color:Word Test: Influence of Age, Sex, and Education; and Normative Data for a Large Sample Across the Adult Age Range. Assessment 13: 62-79. doi: 10.1177/1073191105283427
|
| [10] |
Hanslmayr S, Pastotter B, Bauml KH, et al. (2008) The Electrophysiological Dynamics of Interference During the Stroop Task. J Cognitive Neurosci 20: 215-225. doi: 10.1162/jocn.2008.20020
|
| [11] | Mead LA, Mayer AR, Bobholz JA, et al. (2002) Neural Basis of the Stroop Interference Task: Response Competition or Selective Attention? J Int Neuropsycholo Soc. |
| [12] |
Salo R, Henik A, Robertson LC (2001) Interpreting Stroop Interference: An Analysis of Differences Between Task Versions. Neuropsychology 15: 462-471. doi: 10.1037/0894-4105.15.4.462
|
| [13] | Spieler DH, Balota DA, Faust ME (1996) Stroop Performance in Healthy Younger and Older Adults and in Individuals With Dementia of the Alzheimer's Type. J Experiment Psychol 22: 461-479. |
| [14] |
Thayer JF, Hansen AL, Saus-Rose E, et al. (2009) Heart Rate Variability, Prefrontal Neural Function and Cognitive Performance: the Neurovisceral Integration Perspective on Self:Regulation, Adaptation, and Health. Ann Behav Med 37: 141-53. doi: 10.1007/s12160-009-9101-z
|
| [15] |
Knyazev GG (2007) Motivation, Emotion, and Their Inhibitory Control Mirrored in Brain Oscillations. Neurosci Biobehavior Rev 31: 377-395. doi: 10.1016/j.neubiorev.2006.10.004
|
| [16] |
Alvarez JA, Emory E (2006) Executive Function and the Frontal Lobes: A Meta-Analytic Review. Neuropsychol Rev 16: 17-42. doi: 10.1007/s11065-006-9002-x
|
| [17] |
Botvinick MM, Cohen JD, Carter CS (2004) Conflict Monitoring and Anterior Cingulate Cortex: an Update. Trends Cognit Sci 8: 539-546. doi: 10.1016/j.tics.2004.10.003
|
| [18] | Smith EE, Kosslyn SM (2006) Executive Processes. In: Smith EE, Kosslyn SM, editors. Cognitive Psychology: Mind and Brain. New Jersey: Prentice Hall. pp. 103-146. |
| [19] | Ghimire N, Paudel BH, Khadka R, et al. (2015) Electroencephalographic Changes During Selective Attention. Asian J Med Sci 6: 51-56. |
| [20] |
Milham MP, Erickson KI, Banich MT, et al. (2002) Attentional Control in the Aging Brain: Insights from an fMRI Study of the Stroop Task. Brain Cognition 49: 277-296. doi: 10.1006/brcg.2001.1501
|
| [21] |
Ishii R, Shinosaki K, Ukai S, et al. (1999) Medial Prefrontal Cortex Generates Frontal Midline Theta Rythm. NeuroReport 10: 675-679. doi: 10.1097/00001756-199903170-00003
|
| [22] | Bansal D, Khan M, Salhan AK (2009) A Review of Measurement and Analysis of Heart Rate Variability. Proceedings of the 2009 International Conference on Computer and Automation Engineering: IEEE Computer Society. pp. 243-246. |
| [23] |
Prinsloo GE, Rauch HGL, Karpul D, et al. (2013) The Effect of a Single Session of Short Duration Heart Rate Variability Biofeedback on EEG: A Pilot Study. Appl Psychophysiol Biofeedback 38: 45-56. doi: 10.1007/s10484-012-9207-0
|
| [24] |
Delaney JPA, Brodie DA (2000) Effects of Short:Term Psychological Stress on the Time and Frequency Domains of Heart Rate Variability. Perceptual Motor Skills 91: 515-524. doi: 10.2466/pms.2000.91.2.515
|
| [25] | Salahuddin L (2007) Hear Rate Variability Analysis for Mental Stress Measument in Mobile Settings [Thesis]. Korea: Information and Communications University. pp. 126. |
| [26] |
Dupuy O, Lussier M, Fraser S, et al. (2014) Effect of Overreaching on Cognitive Performance and Related Cardiac Autonomic Control. Scand J Med Sci Sports 24: 234-242. doi: 10.1111/j.1600-0838.2012.01465.x
|
| [27] |
Hovland A, Pallesen S, Hammar A, et al. (2012) The Relationships Among Heart Rate Variability, Executive Functions, and Clinical Variables in Patients With Panic Disorders. Int J Psychophysiol 86: 269-275. doi: 10.1016/j.ijpsycho.2012.10.004
|
| [28] | Petkar HC (2011) Effects of Working Memory Demand on Performance and Mental Stress During the Stroop Task [Thesis]. Canada: Concordia University. pp. 89. |
| [29] |
Oldfield RC (1971) The Assessment and Analysis of Handedness: the Edinburgh Inventory. Neuropsychologia 9: 97-113. doi: 10.1016/0028-3932(71)90067-4
|
| [30] | Klem GH, Lüders HO, Jasper HH, et al. (1999) The Ten:Twenty Electrode System of the International Federation. The International Federation of Clinical Neurophysiology. Int Federation Clinical Neurophysiol Suppl 52: 3-6. |
| [31] | Garcia A, Uribe CE, Tavares MCH, et al. (2011) EEG and Autonomic Responses During Performance of Matching and Non:Matching to Sample Working Memory Tasks With Emotional Content. Front Behavior Neurosci 5: 1-9. |
| [32] |
Delorme A, Makeig S (2004) EEGLAB: an Open Source Toolbox for Analyis of Single:Trial EEG Dynamics Including Independent Component Analysis. J Neurosci Methods 134: 9-21. doi: 10.1016/j.jneumeth.2003.10.009
|
| [33] |
Bell AJ, Sejnowski TJ (1995) An Information:Maximization Approach to Blind Separation and Blind Deconvolution. Neural Comput 7: 1129-1159. doi: 10.1162/neco.1995.7.6.1129
|
| [34] | Belham FS, Satler C, Garcia A, et al. (2013) Age:Related Differences in Cortical Activity During a Visuo:Spatial Working Memory Task with Facial Stimuli. PLOS One 8: 1-8. |
| [35] | Ocazionez SAC (2009) Processamento no Domínio da Frequência de Sinais de Eletroencefalografia Coletados Durante Protocolo de Estresse Moderado. [Dissertação]. Brasília, Distrito Federal: Universidade de Brasília. pp. 119. |
| [36] |
Task Force of the European Society of Cardiology and the North American Society of Pacing and Electrophysiology (1996) Heart Rate Variability: Standards of Measurement, Physiological Interpretation and Clinical Use. Circulation 93: 1043-1065. doi: 10.1161/01.CIR.93.5.1043
|
| [37] | Strauss E, Sherman EMS, Spreen O (2006) Executive Functions. A Compendium of Neuropsychological Tests: Administration, Norms, and Commentary, Third Edition. New York: Oxford University Press. pp. 401-545. |
| [38] |
Klimesch W (2012) Alpha:Band Oscillations, Attention, and Controlled Access to Stored Information. Trends Cognitive Sci 16: 606-617. doi: 10.1016/j.tics.2012.10.007
|
| [39] |
Sauseng P, Klimesch W, Schabus M, et al. (2005) Fronto:parietal EGG Coherence in Theta and Upper Alpha Reflect Central Executive Functions of Working Memory. Int J Psychophysiol 57: 97-103. doi: 10.1016/j.ijpsycho.2005.03.018
|
| [40] | Serrien DJ, Ivry RB, Swinnen SP (2006) Dynamics of Hemispheric Specialization and Integration in the Context of Motor Control. Nat Rev 7: 160-167. |
| [41] | Lent R (2010) Cem Bilhões de Neurônios. São Paulo: Atheneu. pp. 765. |
| [42] |
Boggio PS, Bermpohl F, Vergara AO, et al. (2007) Go:No:Go Task Performance Improvement After Anodal Transcranial Dc Stimulation of the Left Dorsolateral Prefrontal Cortex in Major Depression. J Affect Disorder 101: 91-98. doi: 10.1016/j.jad.2006.10.026
|
| [43] | Knyazev GG (2010) Antero:Posterior EEG Spectral Power Gradient as a Correlate of Extraversion and Behavioural Inhibition. Open Neuroimaging J 4: 114-120. |
| [44] |
Herwig U, Satrapi P, Schonfeldt-Lecuona C (2003) Using the International 10:20 EEG System for Positioning of Transcranial Magnetic Stimulation. Brain Topography 16: 95-99. doi: 10.1023/B:BRAT.0000006333.93597.9d
|
| [45] | Cappa SF, Perani D (2002) Nouns and Verbs: Neurological Correlates of Linguistic Processing. Rivista di Linguistica 14: 73-83. |
| [46] |
Martin A, Haxby JV, Lalonde FM, et al. (1995) Discrete Cortical Regions Associated with Knowledge of Color and Knowledge of Action. Science 270: 102-105. doi: 10.1126/science.270.5233.102
|
| [47] |
Hansen AL, Johnsen BH, Thayer JF (2003) Vagal Influence on Working Memory and Attetion. Int J Psychophysiol 48: 263-274. doi: 10.1016/S0167-8760(03)00073-4
|
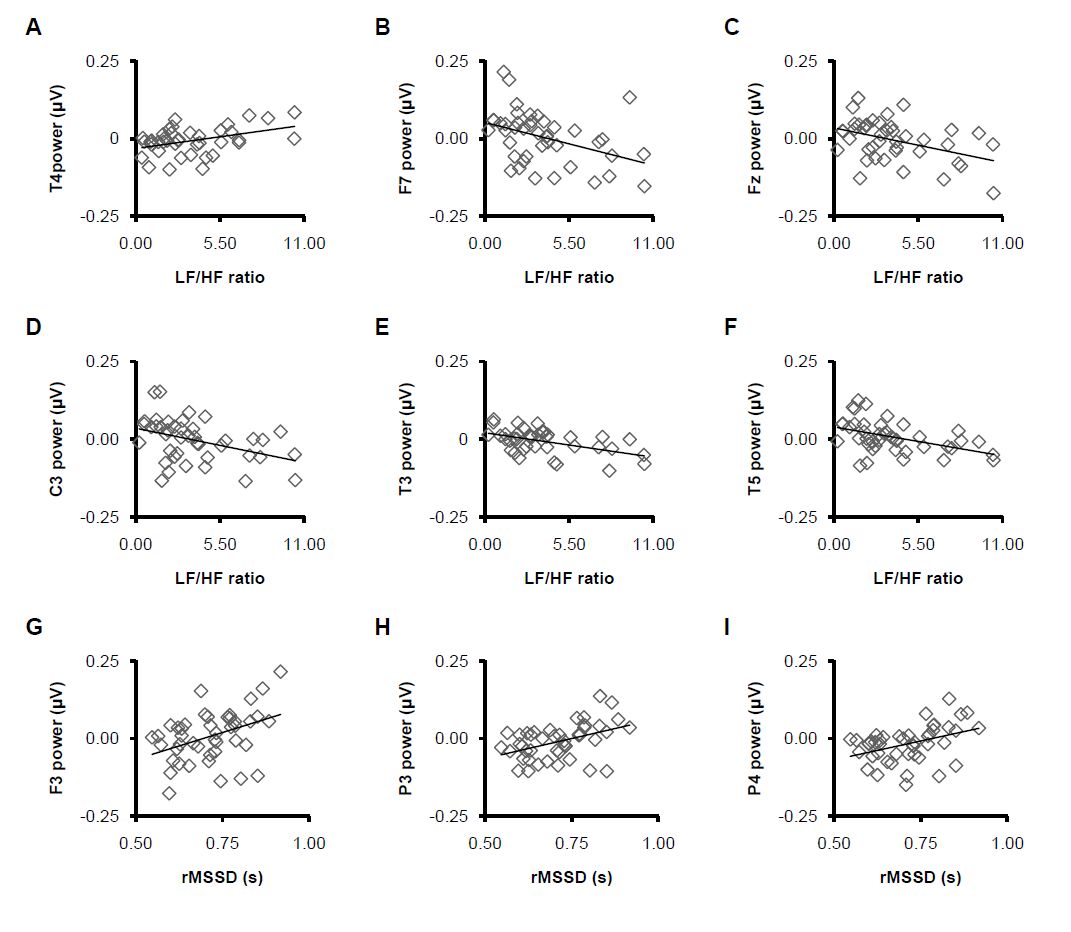
Figures(5) / Tables(2)

Soraya L. Sá Canabarro, Ana Garcia, Corina Satler, Maria Clotilde Henriques Tavares. Interaction between Neural and Cardiac Systems during the Execution of the Stroop Task by Young Adults: Electroencephalographic Activity and Heart Rate Variability[J]. AIMS Neuroscience, 2017, 4(1): 28-51. doi: 10.3934/Neuroscience.2017.1.28







 DownLoad:
DownLoad: